
Sensory Systems/Visual Color Perception
Αντίληψη χρωμάτων
[επεξεργασία]Εισαγωγή
[επεξεργασία]Οι άνθρωποι (μαζί με τα πρωτεύοντα όπως οι μαϊμούδες και οι γορίλες) έχουν την καλύτερη αντίληψη χρώματος μεταξύ των θηλαστικών [1]. Ως εκ τούτου, δεν είναι τυχαίο ότι το χρώμα παίζει σημαντικό ρόλο σε μια ευρεία ποικιλία πτυχών. Για παράδειγμα, το χρώμα είναι χρήσιμο για τη διάκριση και την διαφοροποίηση αντικειμένων, επιφανειών, φυσικών τοπίων, ακόμη και προσώπων [2],[3]. Το χρώμα είναι επίσης ένα σημαντικό εργαλείο για τη μη λεκτική επικοινωνία, συμπεριλαμβανομένου του συναισθήματος [4].
Για πολλές δεκαετίες, ήταν μια πρόκληση να βρεθούν οι δεσμοί μεταξύ των φυσικών ιδιοτήτων του χρώματος και των αντιληπτικών ιδιοτήτων του. Συνήθως, αυτά μελετώνται με δύο διαφορετικές προσεγγίσεις: τη συμπεριφορική απόκριση που προκαλείται από το χρώμα (ονομάζεται επίσης ψυχοφυσική) και την πραγματική φυσιολογική απόκριση που προκαλείται από αυτήν [5].
Εδώ θα επικεντρωθούμε μόνο στην τελευταία. Η μελέτη της φυσιολογικής βάσης της έγχρωμης όρασης, για την οποία σχεδόν τίποτα δεν ήταν γνωστό πριν από το δεύτερο μισό του εικοστού αιώνα, έχει προχωρήσει αργά και σταθερά από το 1950. Σημαντική πρόοδος έχει σημειωθεί σε πολλούς τομείς, ειδικά σε επίπεδο υποδοχέα. Χάρη στις μεθόδους της μοριακής βιολογίας, ήταν δυνατό να αποκαλυφθούν άγνωστες λεπτομέρειες σχετικά με τη γενετική βάση των κηλίδων. Επιπλέον, όλο και περισσότερες περιοχές του φλοιού έχουν δειχθεί ότι επηρεάζονται από οπτικά ερεθίσματα, αν και η συσχέτιση της αντίληψης χρώματος με τη φυσιολογική δραστηριότητα που εξαρτάται από το μήκος κύματος πέρα από τους υποδοχείς δεν είναι τόσο εύκολο να διακριθεί [6].
Σε αυτό το κεφάλαιο, στοχεύουμε να εξηγήσουμε τα βασικά στοιχεία των διαφορετικών διεργασιών της αντίληψης χρώματος κατά μήκος της οπτικής διαδρομής, από τον αμφιβληστροειδή στο μάτι έως τον οπτικό φλοιό στον εγκέφαλο. Για ανατομικές λεπτομέρειες, ανατρέξτε στο Κεφ. "Ανατομία του οπτικού συστήματος" αυτού του Wikibook.
Χρωματική αντίληψη στη αμφιβληστροειδή
[επεξεργασία]Όλα τα χρώματα που μπορούν να διακριθούν από τον άνθρωπο μπορούν να παραχθούν από το μίγμα μόνο τριών βασικών (βασικών) χρωμάτων. Εμπνευσμένο από αυτή την ιδέα της ανάμειξης χρωμάτων, έχει προταθεί ότι το χρώμα υποστηρίζεται από τρεις κατηγορίες αισθητήρων, καθεμία από τις οποίες έχει μέγιστη ευαισθησία σε διαφορετικό τμήμα του ορατού φάσματος [1]. Αρχικά προτάθηκε ρητά το 1853 ότι υπάρχουν τρεις βαθμοί ελευθερίας στη φυσιολογική αντιστοίχιση χρωμάτων [7]. Αυτό επιβεβαιώθηκε αργότερα το 1886 [8] (με αξιοσημείωτα στενά αποτελέσματα σε πρόσφατες μελέτες [9], [10]). Αυτοί οι προτεινόμενοι αισθητήρες χρώματος είναι στην πραγματικότητα οι λεγόμενοι κώνοι (Σημείωση: Σε αυτό το κεφάλαιο, θα ασχοληθούμε μόνο με τους κώνους.) Οι ραβδία συμβάλλουν στην όραση μόνο σε χαμηλά επίπεδα φωτός Αν και είναι γνωστό ότι επηρεάζουν την αντίληψη των χρωμάτων, μικρές και μπορούν να αγνοηθούν εδώ.) [11]. Τα κώνοι είναι από τους δύο τύπους κυττάρων φωτοϋποδοχέα που βρίσκονται στον αμφιβληστροειδή, με σημαντική συγκέντρωση αυτών στον φούντα. Ο παρακάτω πίνακας περιλαμβάνει τους τρεις τύπους κελιών κώνου. Αυτά διακρίνονται από διαφορετικούς τύπους χρωστικής ροδοψίνης. Οι αντίστοιχες καμπύλες απορρόφησης φαίνονται στο παρακάτω σχήμα.
| Name | Higher sensitivity to color | Absorption curve peak [nm] |
|---|---|---|
| S, SWS, B | Blue | 420 |
| M, MWS, G | Green | 530 |
| L, LWS, R | Red | 560 |
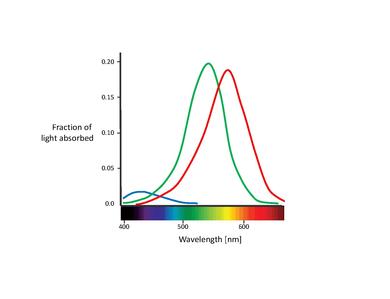
Αν και δεν έχει επιτευχθεί συναίνεση για την ονομασία των διαφορετικών τύπων κώνου, οι πιο ευρέως χρησιμοποιούμενες ονομασίες αναφέρονται είτε στην κορυφή του φάσματος δράσης είτε στο χρώμα στο οποίο είναι ευαίσθητοι (κόκκινο, πράσινο, μπλε) [6]. Σε αυτό το κείμενο, θα χρησιμοποιήσουμε τον ορισμό S-M-L (για βραχύ, μεσαίο και μακρύ μήκος κύματος), αφού αυτά τα ονόματα είναι καταλληλότερα περιγραφικά. Η μπλε-πράσινη-κόκκινη ονοματολογία είναι κάπως παραπλανητική, αφού όλοι οι τύποι κώνων είναι ευαίσθητοι σε ένα μεγάλο εύρος μηκών κύματος.
Ένα σημαντικό χαρακτηριστικό σχετικά με τους τρεις τύπους κώνου είναι η σχετική τους κατανομή στον αμφιβληστροειδή. Αποδεικνύεται ότι οι S-κώνες παρουσιάζουν σχετικά χαμηλή συγκέντρωση μέσω του αμφιβληστροειδούς, λείπουν εντελώς στην πιο κεντρική περιοχή του φούντα. Στην πραγματικότητα, είναι πολύ ευρύχωρα ώστε να διαδραματίζουν σημαντικό ρόλο στην χωρική όραση, παρόλο που είναι ικανά να διαμεσολαβούν στην αδύναμη συνοριακή αντίληψη [12]. Η φόβα κυριαρχείται από L- και M-κώνους. Η αναλογία των δύο τελευταίων μετριέται συνήθως ως λόγος. Έχουν αναφερθεί διαφορετικές τιμές για τον λόγο L / M, που κυμαίνονται από 0,67 [13] έως 2 [14], με τις τελευταίες να είναι οι πλέον αποδεκτές. Γιατί οι L-cones σχεδόν πάντα ξεπερνούν τους M-κώνους παραμένει ασαφής. Παραδόξως, ο λόγος σχετικού κώνου δεν έχει σχεδόν καμία σημαντική επίδραση στην έγχρωμη όραση. Αυτό δείχνει σαφώς ότι ο εγκέφαλος είναι πλαστικός, ικανός να αποκτήσει νόημα από οποιαδήποτε σήματα κώνου λαμβάνει [15], [16].
Είναι επίσης σημαντικό να σημειωθεί η αλληλεπικάλυψη των φασμάτων απορρόφησης L- και M-κώνου. Ενώ το φάσμα απορρόφησης S-κώνου διαχωρίζεται σαφώς, οι κορυφές L- και M-κώνου απέχουν μόνο περίπου 30 nm, ενώ οι φασματικές καμπύλες τους αλληλεπικαλύπτονται επίσης σημαντικά. Αυτό έχει ως αποτέλεσμα μια υψηλή συσχέτιση στα αλιεύματα φωτονίων αυτών των δύο κατηγοριών κώνου. Αυτό εξηγείται από το γεγονός ότι για να επιτευχθεί η μέγιστη δυνατή οξύτητα στο κέντρο της φούντας, το οπτικό σύστημα αντιμετωπίζει τους κώνους L και M εξίσου χωρίς να λαμβάνει υπόψη τα φάσματα απορρόφησης τους. Επομένως, κάθε είδους διαφορά οδηγεί σε υποβάθμιση του σήματος φωτεινότητας [17]. Με άλλα λόγια, ο μικρός διαχωρισμός μεταξύ των φασμάτων L- και M-κώνων μπορεί να ερμηνευθεί ως συμβιβασμός μεταξύ των αναγκών για έγχρωμη όραση υψηλής αντίθεσης και όραση φωτεινότητας υψηλής ακρίβειας. Αυτό είναι σύμφωνο με την έλλειψη S-κώνων στο κεντρικό τμήμα του fovea, όπου η οπτική οξύτητα είναι υψηλότερη. Επιπλέον, η στενή απόσταση των φασμάτων απορρόφησης L- και M-κώνων μπορεί επίσης να εξηγηθεί από τη γενετική τους προέλευση. Και οι δύο τύποι κώνων υποτίθεται ότι έχουν εξελιχθεί "πρόσφατα" (περίπου 35 εκατομμύρια χρόνια πριν) από έναν κοινό πρόγονο, ενώ οι S-κώνοι πιθανώς αποχωρίστηκαν από τον προγονικό παλμό πολύ νωρίτερα [11].
Οι λειτουργίες φασματικής απορρόφησης των τριών διαφορετικών τύπων κωνικών κυττάρων είναι το χαρακτηριστικό γνώρισμα της ανθρώπινης έγχρωμης όρασης. Αυτή η θεωρία επιλύει ένα πολύ γνωστό πρόβλημα: παρόλο που μπορούμε να δούμε εκατομμύρια διαφορετικά χρώματα (οι άνθρωποι μπορούν να διακρίνουν 7 έως 10 εκατομμύρια διαφορετικά χρώματα, οι αμφιβληστροειδοί απλά δεν έχουν αρκετό χώρο για να χωρέσουν μεμονωμένους ανιχνευτές για κάθε χρώμα σε κάθε θέση αμφιβληστροειδούς.
Από τη Ρετίνα στον Εγκέφαλο
[επεξεργασία]Τα σήματα που μεταδίδονται από τον αμφιβληστροειδή σε υψηλότερα επίπεδα δεν είναι απλές σημειακές αναπαραστάσεις των σημάτων υποδοχέα, αλλά μάλλον αποτελούνται από εξελιγμένους συνδυασμούς των σημάτων υποδοχέα. Ο στόχος αυτής της ενότητας είναι να παράσχει μια σύντομη περιγραφή των διαδρομών που λαμβάνουν ορισμένες από αυτές τις πληροφορίες.
Αφού η οπτική εικόνα στον αμφιβληστροειδή μεταδιεγείρεται σε χημικά και ηλεκτρικά σήματα στους φωτοϋποδοχείς, τα σήματα με διαμόρφωση πλάτους μετατρέπονται σε παραστάσεις διαμορφωμένες σε συχνότητα στο κυψελιδικό κύτταρο και σε υψηλότερα επίπεδα. Σε αυτά τα νευρικά κύτταρα, το μέγεθος του σήματος αντιπροσωπεύεται από τον αριθμό των τάσεων τάσης ανά δευτερόλεπτο που τροφοδοτούνται από το κύτταρο και όχι από τη διαφορά τάσης κατά μήκος της κυτταρικής μεμβράνης. Προκειμένου να εξηγηθούν και να αναπαρασταθούν οι φυσιολογικές ιδιότητες αυτών των κυττάρων, θα βρούμε πολύ χρήσιμη την έννοια των δεκτικών πεδίων.
Ένα δεκτικό πεδίο είναι μια γραφική αναπαράσταση της περιοχής στο οπτικό πεδίο στο οποίο αποκρίνεται ένα δεδομένο κύτταρο. Επιπλέον, η φύση της απόκρισης τυπικά υποδεικνύεται για διάφορες περιοχές του δεκτικού πεδίου. Για παράδειγμα, μπορούμε να θεωρήσουμε το πεδίο υποδοχής ενός φωτοϋποδοχέα ως μια μικρή κυκλική περιοχή που αντιπροσωπεύει το μέγεθος και τη θέση της ευαισθησίας αυτού του συγκεκριμένου υποδοχέα στο οπτικό πεδίο. Το σχήμα που ακολουθεί δείχνει παραδειγματικά πεδία υποδοχής για γαγγλιακά κύτταρα, τυπικά σε ανταγωνισμό με κέντρο-περιβάλλον. Το αριστερό πεδίο υποδοχής στο σχήμα απεικονίζει μια θετική κεντρική απόκριση (γνωστή ως επί τόπου). Αυτό το είδος απόκρισης παράγεται συνήθως από μια θετική εισροή από ένα μόνο κώνο που περιβάλλεται από αρνητική απόκριση που παράγεται από πολλούς γειτονικούς κώνους. Επομένως, η απόκριση αυτού του κυττάρου γαγγλίου θα αποτελείται από εισροές από διάφορους κώνους με θετικά και αρνητικά σημεία. Με αυτό τον τρόπο, η κυψέλη δεν ανταποκρίνεται μόνο σε σημεία φωτός, αλλά χρησιμεύει ως ανιχνευτής άκρης (ή πιο σωστά, σημείο). Κατ 'αναλογία με την ορολογία των υπολογιστικών οραμάτων, μπορούμε να σκεφτούμε τις αποκρίσεις κυττάρων γαγγλίου ως αποτέλεσμα μιας συνέλιξης με έναν πυρήνα ανιχνευτή άκρων. Το δεξιό πεδίο υποδοχής στο σχήμα απεικονίζει μια αρνητική κεντρική απόκριση (γνωστή ως off-center), η οποία είναι εξίσου πιθανή. Συνήθως, τα κύτταρα στο κέντρο και εκτός κέντρου θα εμφανιστούν στην ίδια χωρική θέση, τροφοδοτούμενα από τους ίδιους φωτοϋποδοχείς, με αποτέλεσμα μια ενισχυμένη δυναμική εμβέλεια.
Το κατώτερο σχήμα δείχνει ότι εκτός από τον χωρικό ανταγωνισμό, τα γαγγλιακά κύτταρα μπορούν επίσης να έχουν φασματική αντιπαράθεση. Για παράδειγμα, το αριστερό μέρος του κατώτερου σχήματος απεικονίζει μια κόκκινη-πράσινη ανταπόκριση του αντιπάλου με το κέντρο που τροφοδοτείται με θετική είσοδο από έναν L-κώνο και το περιβάλλον που τροφοδοτείται από μια αρνητική είσοδο από Μ-κώνους. Από την άλλη πλευρά, το δεξιό μέρος του κατώτερου σχήματος απεικονίζει την εκτός κέντρου έκδοση αυτής της κελιά. Ως εκ τούτου, προτού οι οπτικές πληροφορίες έχουν βγει από τον αμφιβληστροειδή, έχει ήδη συμβεί επεξεργασία, με βαθιά επίδραση στην εμφάνιση χρώματος. Υπάρχουν και άλλοι τύποι και ποικιλίες απαντήσεων κυττάρων γαγγλίου, αλλά όλες μοιράζονται αυτές τις βασικές έννοιες.
Στο δρόμο προς τον πρωτογενή οπτικό φλοιό, οι άξονες των γαγγλιακών κυττάρων συγκεντρώνονται για να σχηματίσουν το οπτικό νεύρο, το οποίο προβάλλει στον πλευρικό πυρήνα γονιδιώματος (LGN) στον θάλαμο. Η κωδικοποίηση στο οπτικό νεύρο είναι εξαιρετικά αποτελεσματική, διατηρώντας τον αριθμό των νευρικών ινών στο ελάχιστο (περιοριζόμενο από το μέγεθος του οπτικού νεύρου) και έτσι το μέγεθος του τυφλού σημείου του αμφιβληστροειδούς όσο το δυνατόν μικρότερο (περίπου 5 ° πλάτος κατά 7 ° υψηλός). Επιπλέον, τα παρουσιαζόμενα κύτταρα γαγγλίων δεν θα έχουν καμία απάντηση σε ομοιόμορφο φωτισμό, καθώς οι θετικές και οι αρνητικές περιοχές είναι ισορροπημένες. Με άλλα λόγια, τα μεταδιδόμενα σήματα δεν είναι συνδεδεμένα. Για παράδειγμα, οι πληροφορίες από τα γειτονικά τμήματα των φυσικών σκηνών είναι πολύ συσχετισμένες χωρικά και συνεπώς εξαιρετικά προβλέψιμες [18]. Η πλευρική αναστολή μεταξύ γειτονικών γαγγλιοκυττάρων αμφιβληστροειδούς ελαχιστοποιεί αυτή τη χωρική συσχέτιση, βελτιώνοντας έτσι την αποτελεσματικότητα. Μπορούμε να το δούμε ως διαδικασία συμπίεσης εικόνων που πραγματοποιείται στον αμφιβληστροειδή χιτώνα.
Δεδομένης της αλληλεπικάλυψης των φασμάτων απορρόφησης L- και M-κώνου, τα σήματα τους είναι επίσης πολύ συσχετισμένα. Σε αυτή την περίπτωση, η αποτελεσματικότητα της κωδικοποίησης βελτιώνεται συνδυάζοντας τα σήματα κώνου προκειμένου να ελαχιστοποιηθεί η εν λόγω συσχέτιση. Μπορούμε να το κατανοήσουμε πιο εύκολα χρησιμοποιώντας την κύρια ανάλυση στοιχείων (PCA). Το PCA είναι μια στατιστική μέθοδος που χρησιμοποιείται για να μειώσει τη διαστατικότητα ενός δεδομένου συνόλου μεταβλητών μετασχηματίζοντας τις αρχικές μεταβλητές σε ένα σύνολο νέων μεταβλητών, τα κύρια συστατικά (PCs). Ο πρώτος υπολογιστής αντιπροσωπεύει ένα μέγιστο ποσό συνολικής διακύμανσης στις αρχικές μεταβλητές, ο δεύτερος υπολογιστής αντιστοιχεί σε ένα μέγιστο ποσό διακύμανσης που δεν υπολογίστηκε από το πρώτο στοιχείο και ούτω καθεξής. Επιπλέον, οι υπολογιστές είναι γραμμικώς ανεξάρτητοι και ορθογώνιοι ο ένας στον άλλο στο χώρο των παραμέτρων. Το βασικό πλεονέκτημα της PCA είναι ότι μόνο μερικοί από τους ισχυρότερους υπολογιστές είναι αρκετοί για να καλύψουν τη συντριπτική πλειοψηφία της μεταβλητότητας του συστήματος [19]. Αυτό το σχήμα έχει χρησιμοποιηθεί με τις λειτουργίες απορρόφησης κώνου [20] και ακόμη και με τα φυσικώς απαντώμενα φάσματα [21],[22]. Οι υπολογιστές που βρέθηκαν στο χώρο των κωνικών διεγέρσεων που παράγονται από φυσικά αντικείμενα είναι 1) ένας άξονας φωτεινότητας όπου προστίθενται τα σήματα L- και M-κώνων (L + M), 2) η διαφορά του L- και Mκώνου (LM), και 3) έναν άξονα χρώματος όπου το σήμα S-κώνου διαφοροποιείται με το άθροισμα των σημάτων L- και M-κώνων (S- (L + M)). Αυτά τα κανάλια, που προέρχονται από μια μαθηματική / υπολογιστική προσέγγιση, συμπίπτουν με τους τρεις δίαυλους αμφιβληστροειδούς που ανευρίσκονται σε ηλεκτροφυσιολογικά πειράματα [23],[24]. Χρησιμοποιώντας αυτούς τους μηχανισμούς, οι οπτικές περιττές πληροφορίες εξαλείφονται στον αμφιβληστροειδή χιτώνα.
Υπάρχουν τρία κανάλια πληροφοριών που στην πραγματικότητα επικοινωνούν αυτές τις πληροφορίες από τον αμφιβληστροειδή μέσω των κυττάρων των γαγγλίων στο LGN. Διαφέρουν όχι μόνο στις χρωματικές ιδιότητές τους, αλλά και στο ανατομικό τους υπόστρωμα. Αυτά τα κανάλια δημιουργούν σημαντικούς περιορισμούς για βασικές εργασίες χρωμάτων, όπως ανίχνευση και διάκριση.
Στο πρώτο κανάλι, η έξοδος των L- και M-κώνων μεταδίδεται συνεργιστικά σε διάχυτα διπολικά κύτταρα και στη συνέχεια σε κύτταρα στα μαγνητικά στρώματα (Μ-) του LGN (δεν πρέπει να συγχέεται με τους Μ-κώνους του αμφιβληστροειδούς) [25]. Τα δεκτικά πεδία των Μ-κυττάρων αποτελούνται από ένα κέντρο και ένα περιβάλλον, τα οποία είναι χωρικά ανταγωνιστικά. Τα Μκύτταρα έχουν ευαισθησία υψηλής αντίθεσης για τα ερεθίσματα φωταύγειας, αλλά δεν δείχνουν απόκριση σε κάποιο συνδυασμό των εισροών του αντιπάλου LM [26].
Ωστόσο, επειδή τα μηδενικά σημεία διαφορετικών κυττάρων Μ ποικίλλουν ελαφρώς, η αντίδραση του πληθυσμού δεν είναι ποτέ πραγματικά μηδενική. Αυτή η ιδιότητα μεταφέρεται πραγματικά σε φλοιώδεις περιοχές με κυρίαρχη είσοδο κυψελών Μ [26].
Η μονοκυτταρική οδός (Ρ-) προέρχεται από τις μεμονωμένες εξόδους από L- ή Μ-κώνο σε διπολικά κύτταρα του μαστού. Αυτές παρέχουν είσοδο σε Ρ-κύτταρα του αμφιβληστροειδούς [11]. Στον φούντα, τα κέντρα δεκτικών πεδίων των κυττάρων Ρ σχηματίζονται από απλούς L- ή Μ-κώνους. Η δομή του περιβάλλουμενου πεδίου του P-κυττάρου εξακολουθεί να συζητείται. Ωστόσο, η πιο αποδεκτή θεωρία δηλώνει ότι το περίγραμμα αποτελείται από έναν ειδικό τύπο κώνου, καταλήγοντας σε ένα πεδίο υποδοχής του χώρου αντιπάλου για ερεθίσματα φωταύγειας [27]. Τα μεσοκυτταρικά στρώματα συμβάλλουν με το 80% περίπου των συνολικών προβολών από τον αμφιβληστροειδή στο LGN [28].
Τέλος, η πρόσφατα ανακαλυφθείσα οδοκυτταρική οδός (Κ-) μεταφέρει κυρίως σήματα από S-κώνους [29]. Ομάδες αυτού του τύπου κώνων προβάλλουν ειδικά διπολικά κύτταρα, τα οποία με τη σειρά τους παρέχουν εισροή σε συγκεκριμένα μικρά γαγγλιακά κύτταρα. Αυτά συνήθως δεν είναι χωρικά αντίπαλοι. Οι άξονες των μικρών γαγγλιοκυττάρων προβάλλουν σε λεπτά στρώματα του LGN (δίπλα στα πρωτογενή στρώματα) [30].
Ενώ τα κύτταρα των γαγγλιοκυττάρων καταλήγουν στο LGN (δημιουργώντας συνάψεις με κύτταρα LGN), φαίνεται να υπάρχει μία αντιστοιχία μεταξύ των γαγγλιοκυττάρων και των κυττάρων LGN. Το LGN φαίνεται να λειτουργεί ως σταθμός αναμετάδοσης για τα σήματα. Ωστόσο, εξυπηρετεί πιθανώς κάποια οπτική λειτουργία, καθώς υπάρχουν νευρικές προβολές από τον φλοιό πίσω στο LGN που θα μπορούσαν να χρησιμεύσουν ως κάποιο είδος μηχανισμού ανάδρασης ή προσαρμογής. Οι άξονες των κυττάρων LGN προβάλλουν στην οπτική περιοχή ένα (V1) στον οπτικό φλοιό στον ινιακό λοβό.
Χρωματική αντίληψη στον εγκέφαλο
[επεξεργασία]Στον φλοιό, οι προεξοχές από τις μαγνητικές, παρβο- και κυνοκυτταρικές οδούς καταλήγουν σε διαφορετικά στρώματα του πρωτογενούς οπτικού φλοιού. Οι μαγνητοκυτταρικές ίνες νευρώνουν κυρίως το στρώμα 4Cα και το στρώμα 6. Οι παραβολοειδείς νευρώνες προεξέχουν ως επί το πλείστον σε 4Cβ και στα στρώματα 4Α και 6. Οι κωνοκυτταρικοί νευρώνες τερματίζουν στις πλούσιες σε κυτόχρωμα οξειδάσες (CO) στα στρώματα 1, 2 και 3 [31].
Μόλις βρεθεί στον οπτικό φλοιό, η κωδικοποίηση των οπτικών πληροφοριών γίνεται σημαντικά πιο περίπλοκη. Με τον ίδιο τρόπο τα αποτελέσματα των διαφόρων φωτοϋποδοχέων συνδυάζονται και συγκρίνονται με την παραγωγή αποκρίσεων κυττάρων γαγγλίου, τα αποτελέσματα διαφόρων κυττάρων LGN συγκρίνονται και συνδυάζονται για την παραγωγή φλοιώδους αποκρίσεως. Καθώς τα σήματα προχωρούν περισσότερο στην αλυσίδα επεξεργασίας των φλοιών, αυτή η διαδικασία επαναλαμβάνεται με ένα ταχέως αυξανόμενο επίπεδο πολυπλοκότητας μέχρι το σημείο που τα δεκτικά πεδία αρχίζουν να χάνουν νόημα. Ωστόσο, ορισμένες λειτουργίες και διαδικασίες έχουν εντοπιστεί και μελετηθεί σε συγκεκριμένες περιοχές του οπτικού φλοιού.
Στην περιοχή V1 (πτερυγικός φλοιός), οι διπλοί αντίπαλοι νευρώνες - οι νευρώνες που έχουν τα δεκτικοί τους τομείς τόσο χρωματικά όσο και χωρικά αντίθετοι σε σχέση με τις περιοχές on / off ενός μόνο δεκτικού πεδίου - συγκρίνουν τα έγχρωμα σήματα στον οπτικό χώρο [32]. Αποτελούν μεταξύ 5 και 10% των κυττάρων στο V1. Το χονδροειδές μέγεθος και το μικρό τους ποσοστό ταιριάζουν με την κακή χωρική ανάλυση της έγχρωμης όρασης [1]. Επιπλέον, δεν είναι ευαίσθητα στην κατεύθυνση των κινούμενων ερεθισμάτων (σε αντίθεση με κάποιους άλλους νευρώνες V1) και, ως εκ τούτου, είναι απίθανο να συνεισφέρουν στην αντίληψη κίνησης [33]. Ωστόσο, δεδομένης της εξειδικευμένης υποδοχής πεδίου δομή τους, αυτά τα είδη των κυττάρων είναι η νευρωνική βάση για τα έγχρωμα αποτελέσματα αντίθεσης, καθώς και ένα αποτελεσματικό μέσο για την κωδικοποίηση του ίδιου του χρώματος [34], [35]. Άλλα κύτταρα V1 ανταποκρίνονται σε άλλους τύπους ερεθισμάτων, όπως προσανατολισμένες ακμές, διάφορες χωρικές και χρονικές συχνότητες, συγκεκριμένες χωρικές τοποθεσίες και συνδυασμοί αυτών των χαρακτηριστικών, μεταξύ άλλων. Επιπλέον, μπορούμε να βρούμε κύτταρα που συνδυάζουν γραμμικά εισροές από κύτταρα LGN καθώς και κύτταρα που εκτελούν μη γραμμικό συνδυασμό. Αυτές οι απαντήσεις είναι απαραίτητες για την υποστήριξη προηγμένων οπτικών δυνατοτήτων, όπως το ίδιο το χρώμα.

Υπάρχουν σημαντικά λιγότερες πληροφορίες σχετικά με τις χρωματικές ιδιότητες των απλών νευρώνων στο V2 σε σύγκριση με το V1. Από μια πρώτη ματιά, φαίνεται ότι δεν υπάρχουν σημαντικές διαφορές στην κωδικοποίηση των χρωμάτων στα V1 και V2 [36]. Μια εξαίρεση από αυτό είναι η εμφάνιση μιας νέας τάξης κυτταρικού χρώματος κυττάρου [37]. Επομένως, έχει προταθεί ότι η περιοχή V2 συμμετέχει στην επεξεργασία της απόχρωσης. Ωστόσο, αυτό εξακολουθεί να είναι πολύ αμφιλεγόμενο και δεν έχει επιβεβαιωθεί.
Ακολουθώντας την αρθρωτή έννοια που αναπτύχθηκε μετά την ανακάλυψη της λειτουργικής οφθαλμολογικής δεσπόζουσας θέσης στο V1 και λαμβάνοντας υπόψη τον ανατομικό διαχωρισμό μεταξύ των οδών P, M και K (που περιγράφονται στο κεφάλαιο 3), προτάθηκε ότι ένα εξειδικευμένο σύστημα μέσα στο οπτικό πρέπει να υπάρχει ο φλοιός που είναι αφιερωμένος στην ανάλυση των πληροφοριών χρώματος [38]. Το V4 είναι η περιοχή που έχει προσελκύσει ιστορικά την περισσότερη προσοχή ως την πιθανή "περιοχή χρώματος" του εγκεφάλου. Αυτό συμβαίνει λόγω μιας επιρροής μελέτης που ισχυρίστηκε ότι το V4 περιείχε 100% των εκλεκτικών χρωστικών [39]. Ωστόσο, ο ισχυρισμός αυτός αμφισβητήθηκε από αρκετές μετέπειτα μελέτες, μερικοί από τους οποίους ανέφεραν ότι μόνο το 16% των νευρώνων V4 παρουσιάζουν συντονισμό απόχρωσης [40]. Επί του παρόντος, η πιο αποδεκτή ιδέα είναι ότι το V4 συμβάλλει όχι μόνο στο χρώμα, αλλά και στη διαμόρφωση της αντίληψης, της οπτικής προσοχής και της στερεοτυπίας. Επιπλέον, πρόσφατες μελέτες έχουν επικεντρωθεί σε άλλες περιοχές του εγκεφάλου που προσπαθούν να βρουν την "περιοχή χρωμάτων" του εγκεφάλου, όπως το TEO [41] και το PITd [42]. Η σχέση αυτών των περιφερειών μεταξύ τους εξακολουθεί να συζητείται. Για να συμβιβαστεί η συζήτηση, κάποιοι χρησιμοποιούν τον όρο οπίσθιο κατώτερο χρονικό (PIT) φλοιό για να υποδηλώσουν την περιοχή που περιλαμβάνει τα V4, TEO και PITd [1].
Εάν η φλοιώδης απόκριση στα κύτταρα V1, V2 και V4 είναι ήδη πολύ περίπλοκη, το επίπεδο πολυπλοκότητας των σύνθετων οπτικών αποκρίσεων σε ένα δίκτυο περίπου 30 οπτικών ζωνών είναι τεράστιο. Το σχήμα 4 δείχνει ένα μικρό τμήμα της συνδεσιμότητας των διαφορετικών φλοιωδών περιοχών (όχι των κυττάρων) που έχουν ταυτοποιηθεί [43].
Σε αυτό το στάδιο, γίνεται εξαιρετικά δύσκολο να εξηγηθεί η λειτουργία των κυττάρων μεμονωμένων φλοιών με απλούς όρους. Στην πραγματικότητα, η λειτουργία ενός μόνο κυττάρου μπορεί να μην έχει νόημα, αφού η αναπαράσταση διαφόρων αντιλήψεων πρέπει να κατανέμεται σε συλλογές κυττάρων σε ολόκληρο τον φλοιό.
Μηχανισμοί προσαρμογής χρωμάτων
[επεξεργασία]Αν και οι ερευνητές προσπαθούν να εξηγήσουν την επεξεργασία των σημάτων χρώματος στο ανθρώπινο οπτικό σύστημα, είναι σημαντικό να καταλάβουμε ότι η αντίληψη χρώματος δεν είναι μια σταθερή διαδικασία. Στην πραγματικότητα, υπάρχουν διάφοροι δυναμικοί μηχανισμοί που χρησιμεύουν για τη βελτιστοποίηση της οπτικής απόκρισης ανάλογα με το περιβάλλον προβολής. Ιδιαίτερη σημασία για την αντίληψη χρώματος είναι οι μηχανισμοί σκοτεινής, ελαφριάς και χρωματικής προσαρμογής.
Σκοτεινή προσαρμογή
[επεξεργασία]Η σκοτεινή προσαρμογή αναφέρεται στην αλλαγή στην οπτική ευαισθησία που συμβαίνει όταν μειώνεται το επίπεδο φωτισμού. Η απόκριση του οπτικού συστήματος σε μειωμένο φωτισμό είναι να γίνει πιο ευαίσθητη, αυξάνοντας την ικανότητά του να παράγει μια ουσιαστική οπτική απόκριση ακόμη και όταν οι συνθήκες φωτισμού δεν είναι βέλτιστες [44].
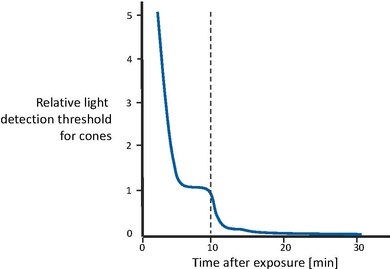
Το σχήμα 5 δείχνει την ανάκτηση της οπτικής ευαισθησίας μετά τη μετάβαση από ένα εξαιρετικά υψηλό επίπεδο φωτισμού σε ένα πλήρες σκοτάδι [43]. Κατ 'αρχάς, οι κώνοι γίνονται σταδιακά πιο ευαίσθητοι, έως ότου η καμπύλη ξεπεράσει μετά από μερικά λεπτά. Στη συνέχεια, μετά από περίπου 10 λεπτά, η οπτική ευαισθησία είναι σχεδόν σταθερή. Σε αυτό το σημείο, το σύστημα ράβδων, με μεγαλύτερο χρόνο ανάκτησης, έχει ανακτήσει αρκετή ευαισθησία ώστε να ξεπεράσει τους κώνους και επομένως να ανακτήσει τον έλεγχο της συνολικής ευαισθησίας. Η ευαισθησία της ραβδίου βελτιώνεται σταδιακά, μέχρι να γίνει ασυμπτωτική μετά από περίπου 30 λεπτά. Με άλλα λόγια, οι κώνοι είναι υπεύθυνοι για την ανάκτηση ευαισθησίας για τα πρώτα 10 λεπτά. Στη συνέχεια, οι ραβδία ξεπερνούν τους κώνους και αποκτούν πλήρη ευαισθησία μετά από περίπου 30 λεπτά.
Αυτός είναι μόνο ένας από τους πολλούς νευρικούς μηχανισμούς που παράγονται προκειμένου να προσαρμόζονται στις συνθήκες σκοτεινής κεραίας όσο το δυνατόν καλύτερο. Κάποιοι άλλοι νευρωνικοί μηχανισμοί περιλαμβάνουν το γνωστό αντανακλαστικό των μαθητών, την εξάντληση και την αναγέννηση της φωτοχρωματογραφίας, τον έλεγχο κέρδους στα κύτταρα του αμφιβληστροειδούς και άλλους μηχανισμούς υψηλότερου επιπέδου και τη γνωστική ερμηνεία μεταξύ άλλων.
Προσαρμογή φωτός
[επεξεργασία]Η προσαρμογή φωτός είναι ουσιαστικά η αντίστροφη διαδικασία της σκοτεινής προσαρμογής. Στην πραγματικότητα, οι βασικοί φυσιολογικοί μηχανισμοί είναι οι ίδιοι και για τις δύο διαδικασίες. Ωστόσο, είναι σημαντικό να το εξετάσουμε χωριστά, καθώς οι οπτικές του ιδιότητες διαφέρουν.

Η προσαρμογή φωτός συμβαίνει όταν αυξάνεται το επίπεδο φωτισμού. Επομένως, το οπτικό σύστημα πρέπει να γίνει λιγότερο ευαίσθητο για να παράγει χρήσιμες αντιλήψεις, δεδομένου ότι υπάρχει πολύ πιο ορατό φως διαθέσιμο. Το οπτικό σύστημα διαθέτει μια περιορισμένη δυναμική περιοχή εξόδου διαθέσιμη για τα σήματα που παράγουν τις αντιλήψεις μας. Ωστόσο, ο πραγματικός κόσμος έχει επίπεδα φωτισμού που καλύπτουν τουλάχιστον 10 τάξεις μεγέθους περισσότερο. Ευτυχώς, σπάνια πρέπει να βλέπουμε όλο το φάσμα των επιπέδων φωτισμού ταυτόχρονα.
Σε υψηλά επίπεδα φωτισμού, η προσαρμογή επιτυγχάνεται με λεύκανση με φωτοχρωστικές ουσίες. Αυτό κλιμακώνει τη δέσμευση φωτονίων στους υποδοχείς και προστατεύει την απόκριση του κώνου από κορεσμό σε φωτεινά υπόβαθρα. Οι μηχανισμοί προσαρμογής φωτός εμφανίζονται κυρίως στον αμφιβληστροειδή [45]. Στην πραγματικότητα, οι μεταβολές κέρδους είναι σε μεγάλο βαθμό κωνικές και τα σήματα προσαρμογής συγκεντρώνονται σε περιοχές που δεν είναι μεγαλύτερες από τη διάμετρο των μεμονωμένων κώνων [46], [47]. Αυτό δείχνει τον εντοπισμό προσαρμογής φωτός που μπορεί να είναι ήδη από τους υποδοχείς. Ωστόσο, φαίνεται ότι υπάρχουν περισσότερες από μία τοποθεσίες κλιμάκωσης ευαισθησίας. Ορισμένες από τις αλλαγές κέρδους είναι εξαιρετικά γρήγορες, ενώ άλλες παίρνουν δευτερόλεπτα ή και λεπτά για να σταθεροποιηθούν [48]. Συνήθως, η προσαρμογή φωτός διαρκεί περίπου 5 λεπτά (έξι φορές ταχύτερα από τη σκοτεινή προσαρμογή). Αυτό μπορεί να δείχνει την επίδραση των μεταδεκτικών περιοχών.
Το σχήμα 6 δείχνει παραδείγματα προσαρμογής φωτός [43]. Εάν χρησιμοποιήσαμε μία μόνο λειτουργία απόκρισης για να χαρτογραφήσουμε το μεγάλο εύρος εντάσεων στην έξοδο του οπτικού συστήματος, τότε θα είχαμε μόνο μια πολύ μικρή κλίμακα στη διάθεσή μας για μια δεδομένη σκηνή. Είναι σαφές ότι με μια τέτοια λειτουργία απόκρισης, η αντιληπτή αντίθεση οποιασδήποτε δεδομένης σκηνής θα ήταν περιορισμένη και η οπτική ευαισθησία στις αλλαγές θα υποβαθμιζόταν σοβαρά λόγω ζητημάτων σήματος-θορύβου. Αυτή η περίπτωση φαίνεται από τη διακεκομμένη γραμμή. Από την άλλη πλευρά, οι συμπαγείς γραμμές αντιπροσωπεύουν οικογένειες οπτικών αποκρίσεων. Αυτές οι καμπύλες καταγράφουν το εύρος χρήσιμου φωτισμού σε οποιαδήποτε δεδομένη σκηνή στο πλήρες δυναμικό εύρος της οπτικής εξόδου, με αποτέλεσμα την καλύτερη οπτική αντίληψη για κάθε κατάσταση. Η προσαρμογή φωτός μπορεί να θεωρηθεί ως η διαδικασία της ολίσθησης της καμπύλης οπτικής απόκρισης κατά μήκος του άξονα της στάθμης φωτισμού έως ότου επιτευχθεί το βέλτιστο επίπεδο για τις δεδομένες συνθήκες προβολής.
Χρωμική προσαρμογή
[επεξεργασία]Η γενική έννοια της χρωματικής προσαρμογής συνίσταται στην μεταβολή του ύψους των καμπυλών φασματικής απόκρισης των τριών κώνων. Αυτή η ρύθμιση προκύπτει επειδή η προσαρμογή φωτός συμβαίνει ανεξάρτητα σε κάθε κατηγορία κώνου. Μια συγκεκριμένη διατύπωση αυτής της υπόθεσης είναι γνωστή ως προσαρμογή von Kries. Αυτή η υπόθεση δηλώνει ότι η απόκριση προσαρμογής λαμβάνει χώρα σε κάθε έναν από τους τρεις τύπους κώνων χωριστά και ισοδυναμεί με τον πολλαπλασιασμό των σταθερών φασματικών ευαισθησιών τους με μία σταθερά κλίμακας [49]. Εάν τα βάρη κλίμακας (επίσης γνωστά ως συντελεστές von Kries) είναι αντιστρόφως ανάλογες με την απορρόφηση του φωτός από κάθε τύπο κώνου (δηλαδή χαμηλότερη απορρόφηση θα απαιτήσει μεγαλύτερο συντελεστή), τότε η κλίμακα von Kries διατηρεί μια σταθερή μέση απόκριση σε κάθε κατηγορία κώνου. Αυτό παρέχει έναν απλό αλλά ισχυρό μηχανισμό για τη διατήρηση του αντιληπτού χρώματος αντικειμένων παρά τις αλλαγές στον φωτισμό. Κάτω από μια σειρά διαφορετικών συνθηκών, η κλίμακα von Kries παρέχει μια καλή εικόνα των επιπτώσεων της προσαρμογής φωτός στην ευαισθησία και την εμφάνιση χρώματος [50], [51].
Ο ευκολότερος τρόπος για να απεικονίσετε τη χρωματική προσαρμογή είναι να εξετάσετε ένα λευκό αντικείμενο υπό διαφορετικούς τύπους φωτισμού. Για παράδειγμα, ας εξετάσουμε την εξέταση ενός τεμαχίου χαρτιού κάτω από το φως της ημέρας, φθορισμού και φωτισμού πυρακτώσεως. Το φως της ημέρας περιέχει σχετικώς πολύ περισσότερη ενέργεια μικρού μήκους κύματος από το φθορίζον φως και ο φωτισμός πυρακτώσεως περιέχει σχετικώς πολύ περισσότερη ενέργεια μακρού μήκους κύματος από το φθορίζον φως. Ωστόσο, παρά τις διαφορετικές συνθήκες φωτισμού, το χαρτί διατηρεί περίπου την λευκή του εμφάνιση κάτω από τις τρεις πηγές φωτός. Αυτό συμβαίνει επειδή το σύστημα S-cone γίνεται σχετικά λιγότερο ευαίσθητο υπό το φως της ημέρας (προκειμένου να αντισταθμιστεί η πρόσθετη ενέργεια μικρού μήκους κύματος) και το σύστημα Lκώνων καθίσταται σχετικά λιγότερο ευαίσθητο υπό τον φωτισμό πυρακτώσεως (προκειμένου να αντισταθμιστεί η πρόσθετη μακροπρόθεσμη ακτινοβολία) ενέργεια μήκους κύματος) [43].
- ↑ Conway, Bevil R (2009). "Color vision, cones, and color-coding in the cortex". The neuroscientist 15: 274-290.
- ↑ Russell, Richard and Sinha, Pawan} (2007). "Real-world face recognition: The importance of surface reflectance properties". Perception 36 (9).
- ↑ Gegenfurtner, Karl R and Rieger, Jochem (2000). "Sensory and cognitive contributions of color to the recognition of natural scenes". Current Biology 10 (13): 805-808.
- ↑ Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke (2006). "Bare skin, blood and the evolution of primate colour vision". Biology letters 2 (2): 217-221.
- ↑ Beretta, Giordano (2000). Understanding Color. Hewlett-Packard.
- ↑ 6,0 6,1 Boynton, Robert M (1988). "Color vision". Annual review of psychology 39 (1): 69-100.
- ↑ Grassmann, Hermann (1853). "Zur theorie der farbenmischung". Annalen der Physik 165 (5): 69-84.
- ↑ Konig, Arthur and Dieterici, Conrad (1886). "Die Grundempfindungen und ihre intensitats-Vertheilung im Spectrum". Koniglich Preussischen Akademie der Wissenschaften.
- ↑ Smith, Vivianne C and Pokorny, Joel (1975). "Spectral sensitivity of the foveal cone photopigments between 400 and 500 nm". Vision research 15 (2): 161-171.
- ↑ Vos, JJ and Walraven, PL (1971). "On the derivation of the foveal receptor primaries". Vision Research 11 (8): 799-818.
- ↑ 11,0 11,1 Gegenfurtner, Karl R and Kiper, Daniel C (2003). "Color vision". Neuroscience 26 (1): 181.
- ↑ Kaiser, Peter K and Boynton, Robert M (1985). "Role of the blue mechanism in wavelength discrimination". Vision research 125 (4): 523-529.
- ↑ Paulus, Walter and Kroger-Paulus, Angelika (1983). "A new concept of retinal colour coding". Vision research 23 (5): 529-540.
- ↑ Nerger, Janice L and Cicerone, Carol M (1992). "The ratio of L cones to M cones in the human parafoveal retina". Vision research 32 (5): 879-888.
- ↑ Neitz, Jay and Carroll, Joseph and Yamauchi, Yasuki and Neitz, Maureen and Williams, David R (2002). "Color perception is mediated by a plastic neural mechanism that is adjustable in adults". Neuron 35 (4): 783-792.
- ↑ Jacobs, Gerald H and Williams, Gary A and Cahill, Hugh and Nathans, Jeremy (2007). "Emergence of novel color vision in mice engineered to express a human cone photopigment". Science 315 (5819): 1723-1725.
- ↑ Osorio, D and Ruderman, DL and Cronin, TW (1998). "Estimation of errors in luminance signals encoded by primate retina resulting from sampling of natural images with red and green cones". JOSA A 15 (1): 16-22.
- ↑ Kersten, Daniel (1987). "Predictability and redundancy of natural images". JOSA A 4 (112): 2395-2400.
- ↑ Jolliffe, I. T. (2002). Principal Component Analysis. Springer.
- ↑ Buchsbaum, Gershon and Gottschalk, A (1983). "Trichromacy, opponent colours coding and optimum colour information transmission in the retina". Proceedings of the Royal society of London. Series B. Biological sciences 220 (1218): 89-113.
- ↑ Zaidi, Qasim (1997). "Decorrelation of L-and M-cone signals". JOSA A 14 (12): 3430-3431.
- ↑ Ruderman, Daniel L and Cronin, Thomas W and Chiao, Chuan-Chin (1998). "Statistics of cone responses to natural images: Implications for visual coding". JOSA A 15 (8): 2036-2045.
- ↑ Lee, BB and Martin, PR and Valberg, A (1998). "The physiological basis of heterochromatic flicker photometry demonstrated in the ganglion cells of the macaque retina". The Journal of Physiology 404 (1): 323-347.
- ↑ Derrington, Andrew M and Krauskopf, John and Lennie, Peter (1984). "Chromatic mechanisms in lateral geniculate nucleus of macaque". The Journal of Physiology 357 (1): 241-265.
- ↑ Martin, Paul R and Lee, Barry B and White, Andrew JR and Solomon, Samuel G and Ruttiger, Lukas (2001). "Chromatic sensitivity of ganglion cells in the peripheral primate retina". Nature 410 (6831): 933-936.
- ↑ Perry, VH and Oehler, R and Cowey, A (1984). "Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey". Neuroscience 12 (4): 1101--1123.