
Sensory Systems/Visual System
Πρότυπο:SensorySystems Navigation
Εισαγωγή
[επεξεργασία]Τα οπτικά συστήματα βασίζονται σε γενικές γραμμἐς σε ηλεκτρομαγνητικά (ΗM) κύματα για να δώσουν στον οργανισμό περισσότερες πληροφορίες γὐρω από το περιβάλλον του. Αυτές οι πληροφορίες θα μπορούσαν να αφορούν πιθανους συντρόφους, κινδύνους και πηγές διατροφής. Οι διαφορετικοί οργανισμοί έχουν διαφορετικά στοιχεία που συνθέτουν αυτό που αναφέρεται ως οπτικό σύστημα.
Η πολυπλοκότητα των ματιών κυμαίνεται από κάτι τόσο απλό όσο μια οπτική κηλίδα, η οποία δεν είναι τίποτα περισσότερο από μια συλλογή φωτοευαίσθητων κυττάρων, ως ένα πλήρως ανεπτυγμένο φακό κάμερας. Εάν ένας οργανισμός έχει διαφορετικούς τύπους φωτοευαίσθητων κυττάρων ή κυττάρων ευαίσθητων σε διαφορετικές περιοχές μήκους κύματος, ο οργανισμός θα μπορούσε θεωρητικά να αντιληφθεί το χρώμα ή τουλάχιστον τις διαφορές χρώματος. Η πόλωση, μια άλλη ιδιότητα της ΗΜ ακτινοβολίας, μπορεί να ανιχνευθεί από ορισμένους οργανισμούς, με έντομα και κεφαλόποδα να εχουν την υψηλότερη ακρίβεια.
Θα πρεπει να σημειωθεί ότι αυτό το κείμενο εστιάζεται στη χρήση των ορατων ΗM κυμάτων. Σιγουρα, ορισμένοι οργανισμοί έχουν αναπτύξει εναλλακτικούς τρόπους για να αποκτήσουν όραση ή τουλάχιστον για να συμπληρώσουν αυτά που βλέπουν με εξω-αισθητηριακές πληροφορίες. Παράδειγμα αποτελουν οι φάλαινες ή οι νυχτερίδες, οι οποίες χρησιμοποιούν ηχώ. Αυτες μπορεί να βλέπουν συμφωνα με τον ορισμό της λέξης, αλλά αυτο δεν είναι απολύτως σωστό. Επιπλέον, όραση και οπτικο είναι λέξεις που συνηθέστερα συνδέονται με τα ΗM κύματα στην περιοχή οπτικού μήκους κύματος, που κανονικά καθορίζεται απο τα ίδια τα όρια του μήκους κύματος της ανθρώπινης όρασης.

Δεδομένου ότι ορισμένοι οργανισμοί ανιχνεύουν τα ΗM κύματα με συχνότητες ανωτερες και
κατωτερες από εκείνες των ανθρώπων, θα πρέπει να δοθει ενας καλύτερος
ορισμός. Επομένως, ορίζουμε την περιοχή οπτικού μήκους κύματος ως το μήκος
ΗΜ κύματος μεταξύ 300nm και 800nm. Αυτό μπορεί να φαίνεται αυθαίρετο σε
μερικούς, αλλά η επιλογή λανθασμένων ορίων θα καθιστούσε τμήματα της ορασης
ενός πουλιού ως μη ορατη. Επίσης, με αυτό το φάσμα μηκους κύματος, έχουμε
ορίσει, για παράδειγμα, τη θερμική όραση ορισμένων οργανισμών, όπως για
παράδειγμα τα φίδια, ως μη όραση. Ως εκ τούτου, τα φίδια που χρησιμοποιούν
τα σωληνωτά όργανά τους, τα οποία είναι ευαίσθητα σε ΗM μεταξύ 5000nm και
30,000nm (IR), δεν "βλέπουν", αλλά κατά κάποιο τρόπο "αισθάνονται" από
μακριά. Ακόμη και αν τυφλά δείγματα έχουν τεκμηριωθεί στοχεύοντας και
προσβάλλοντας συγκεκριμένα μέρη του σώματος.
Αρχικά θα εκπονηθεί μια σύντομη περιγραφή των διαφόρων τύπων αισθητήριων
οργάνων του οπτικού συστήματος, ακολουθούμενη από μια διεξοδική εξήγηση των
συστατικών στην ανθρώπινη όραση, την επεξεργασία σήματος της οπτικής οδού
στον άνθρωπο, στο τἐλος θα δοθεἰ ένα παράδειγμα του αισθητικού
αποτελέσματος που οφείλεται αυτά τα στάδια.
Αισθητήρια όργανα
[επεξεργασία]Η όραση ή απλά η ικανότητα του να βλέπεις, εξαρτάται από τα αισθητήρια όργανα ή τα μάτια του οπτικού συστήματος. Υπάρχουν πολλές διαφορετικές κατασκευές των ματιών, που κυμαίνονται στην πολυπλοκότητα ανάλογα με τις απαιτήσεις του οργανισμού. Οι διαφορετικές κατασκευές έχουν διαφορετικές δυνατότητες, είναι ευαίσθητες σε διαφορετικά μήκη κύματος και έχουν διαφορετικούς βαθμούς οξύτητας, απαιτούν επίσης διαφορετική επεξεργασία ώστε να έχουν νόημα για την είσοδο και διαφορετικούς βαθμούς για να λειτουργούν άριστα. Η ικανότητα ανίχνευσης και αποκρυπτογράφησης του ΗΜ έχει αποδειχθεί πολύτιμο πλεονέκτημα στις περισσότερες μορφές ζωής, οδηγώντας σε αυξημένη πιθανότητα επιβίωσης για τους οργανισμούς που το χρησιμοποιούν. Σε περιβάλλοντα χωρίς επαρκή φωτισμό ή πλήρη έλλειψη αυτοὐ, οι μορφές ζωής δεν έχουν το πλεονέκτημα της όρασης, το οποίο τελικά έχει οδηγήσει σε ατροφία οπτικών αισθητήριων οργάνων με επακόλουθη αυξημένη εξάρτηση από άλλες αισθήσεις (π.χ. ζώα που ζουν σε σπήλαια, νυχτερίδες κλπ.). Είναι ενδιαφέρον το γεγονός ότι τα οπτικά αισθητήρια όργανα συντονίζονται στο οπτικό παράθυρο, το οποίο ορίζεται από τα μήκη κύματος EM (μεταξύ 300nm και 1100nm) που περνούν από την ατμόσφαιρα και φτάνει στο έδαφος. Αυτό φαίνεται στο παρακάτω σχήμα. Μπορεί να παρατηρήσετε ότι υπάρχουν και άλλα παράθυρα, ένα παράθυρο IR, το οποίο εξηγεί σε κάποιο βαθμό η θερμική όραση των φιδιών και ένα παράθυρο ραδιοσυχνοτήτων (RF), το οποίο δεν είναι σε θέση να ανιχνεύσει καμία γνωστή μορφή ζωής. Μέσα στον χρόνο η εξέλιξη δημιούργησε πολλές κατασκευές οφθαλμών και μερικές εξελίχθηκαν πολλές φορές, αποδίδοντας ομοιότητες με οργανισμούς που αποτελλούν παρόμοιες εξαιρέσεις.

Υπάρχει μια πτυχή που είναι ουσιαστικά ταυτόσημη, ανεξάρτητα από το είδος ή την πολυπλοκότητα του αισθητήριου τύπου οργάνου, η καθολική χρήση των ευαίσθητων στο φως πρωτεϊνών που ονομάζονται ”opsins”. Χωρίς να εστιάζουμε πάρα πολύ στη μοριακή βάση, οι διάφορες κατασκευές μπορούν να κατηγοριοποιηθούν σε ξεχωριστές ομάδες:
- Οφθαλμοἰ σημειακἠς εστἰασης
- Οφθαλμοἰ κατεύθυνσης
- Στενοειδεἰς οφθαλμοἰ
- Οφθαλμοἰ φακού
- Διαθλαστικά μάτια κερατοειδούς
- Ανακλαστικοἰ οφθαλμοἰ
- Σύνθετοι οφθαλμοἰ
Η λιγότερο πολὐπλοκη διαμόρφωση των ματιών επιτρέπει στους οργανισμούς να αισθανθούν απλά το φως του περιβάλλοντος, επιτρέποντας στον οργανισμό να γνωρίζει αν υπάρχει φως ή όχι. Συνήθως είναι απλά μια συλλογή φωτοευαίσθητων κυττάρων σε ένα σύμπλεγμα στο ίδιο σημείο, έτσι μερικές φορές αναφέρεται ως μάτια σημείου, σημείο ματιών ή στέλεχος. Με την προσθήκη περισσότερων γωνιακών δομών ή την εσοχή των οφθαλμικών ματιών, ένας οργανισμός αποκτά πρόσβαση και στις κατευθυντήριες πληροφορίες, γεγονός που αποτελεί ζωτική απαίτηση για τον σχηματισμό εικόνας. Αυτά τα αποκαλούμενα μάτια είναι οι πιο συνηθισμένοι τύποι οπτικών αισθητηρίων οργάνων και μπορούν να βρεθούν σε πάνω από το 95% όλων των γνωστών ειδών.
.jpg)
Ξεκινώντας από το προφανές ακραίο αυτής της προσέγγισης, οδηγούμαστε σε μια σπηλαιώδης δομή, η οποία αυξάνει την ευκρίνεια της εικόνας, δυστυχώς με απώλεια έντασης. Με άλλα λόγια, υπάρχει μια ανταλλαγή ανάμεσα στην ένταση ή τη φωτεινότητα και την οξύτητα. Ένα παράδειγμα αυτού μπορεί να βρεθεί στο Nautilus, είδη που ανήκουν στην οικογένεια Nautilidae, οργανισμοί που θεωρούνται ως ζωντανά απολιθώματα. Είναι το μόνο γνωστό είδος που έχει αυτόν τον τύπο ματιού, που αναφέρεται ως το μάτι της οπής, και είναι εντελώς ανάλογο με την κάμερα οπών ή την κάμερα obscura. Επιπλέον, όπως και οι πιο προηγμένες κάμερες, η Nautili μπορεί να προσαρμόσει το μέγεθος του ανοίγματος αυξάνοντας ή μειώνοντας έτσι την ανάλυση του οφθαλμού σε αντίστοιχη μείωση ή αύξηση της φωτεινότητας της εικόνας. Όπως και η κάμερα, ο τρόπος μείωσης του προβλήματος έντασης / ανάλυσης είναι να συμπεριληφθεί ένας φακός, μια δομή που επικεντρώνει το φως σε μια κεντρική περιοχή, η οποία συχνότερα έχει υψηλότερη πυκνότητα φωτοαισθητήρων. Ρυθμίζοντας το σχήμα του φακού και μετακινώντας το γύρω και ελέγχοντας το μέγεθος του ανοίγματος ή της κόρης, οι οργανισμοί μπορούν να προσαρμοστούν σε διαφορετικές συνθήκες και να επικεντρωθούν σε συγκεκριμένες περιοχές ενδιαφέροντος σε οποιαδήποτε οπτική σκηνή. Η τελευταία αναβάθμιση στις διάφορες κατασκευές των ματιών που ήδη αναφέρθηκαν είναι η συμπερίληψη ενός διαθλαστικού κερατοειδούς. Τα μάτια με αυτή τη δομή έχουν μεταβιβάσει τα δύο τρίτα της συνολικής οπτικής ισχύος του οφθαλμού στο υγρό υψηλού δείκτη διάθλασης μέσα στον κερατοειδή, επιτρέποντας την όραση πολύ υψηλής ανάλυσης. Τα περισσότερα χερσαία ζώα, συμπεριλαμβανομένων των ανθρώπων, έχουν τα μάτια αυτής της συγκεκριμένης κατασκευής. Επιπλέον, υπάρχουν πολλές παραλλαγές της δομής του φακού, του αριθμού του φακού, της πυκνότητας του φωτοαισθητήρα, του σχήματος του βοθρίου, του αριθμού του βοθρίου, του σχήματος της κόρης κ.λπ., προκειμένου να αυξηθούν οι πιθανότητες επιβίωσης για τον εν λόγω οργανισμό. Αυτές οι παραλλαγές οδηγούν σε ποικίλη εξωτερική εμφάνιση των ματιών, ακόμη και με μία κατηγορία κατασκευής οφθαλμών. Για να αποδειχθεί αυτό το σημείο, παρουσιάζεται μια συλλογή φωτογραφιών ζώων με την ίδια κατηγορία ματιών (διαθλαστικά μάτια κερατοειδούς).
 |
 |
 |
 |
.jpg) |
Μια εναλλακτική λύση στην προσέγγιση του φακού που ονομάζεται μάτι ανακλαστήρα μπορεί να βρεθεί για παράδειγμα στα μαλάκια. Αντί του συμβατικού τρόπου εστίασης του φωτός σε ένα μόνο σημείο στο πίσω μέρος του ματιού χρησιμοποιώντας έναν φακό ή ένα σύστημα φακών, οι οργανισμοί αυτοί έχουν καθρέφτες σαν δομές στο εσωτερικό του θαλάμου του ματιού που αντανακλούν το φως σε ένα κεντρικό τμήμα, σαν ένα παραβολικό πιάτο. Παρόλο που δεν υπάρχουν γνωστά παραδείγματα οργανισμών με αντανακλαστικά μάτια ικανά για σχηματισμό εικόνας, τουλάχιστον ένα είδος ψαριού, τα Dolichopteryx longipes τα χρησιμοποιούν σε συνδυασμό με τα "κανονικά" φακοειδή μάτια.

Η τελευταία ομάδα οφθαλμών, που βρίσκεται στα έντομα και τα καρκινοειδή, ονομάζεται σύνθετα μάτια. Αυτά τα μάτια αποτελούνται από μια σειρά λειτουργικών υπομονάδων που ονομάζονται ommatidia, το καθένα από τα οποία αποτελείται από μια επιφάνεια ή πρόσθια επιφάνεια, ένα διαφανές κρυσταλλικό κώνο και φωτοευαίσθητα κύτταρα για ανίχνευση. Επιπλέον, κάθε μία από τις ommatidia διαχωρίζεται από κύτταρα χρωστικής, διασφαλίζοντας ότι το εισερχόμενο φως είναι όσο το δυνατόν παράλληλο. Ο συνδυασμός των αποτελεσμάτων καθεμιάς από αυτές τις ommatidia σχηματίζει μια ψηφιδωτή εικόνα, με ένα ψήφισμα ανάλογο του αριθμού των μονάδων ommatidia. Για παράδειγμα, αν οι άνθρωποι είχαν σύνθετα μάτια, τα μάτια θα είχαν καλύψει ολόκληρα τα πρόσωπά μας για να διατηρήσουν το ίδιο ψήφισμα. Σημειωτέον, υπάρχουν πολλοί τύποι σύνθετων οφθαλμών, αλλά η εμβάθυνση σε αυτό το θέμα είναι πέρα από το πεδίο εφαρμογής αυτού του κειμένου.
Όχι μόνο ο τύπος των ματιών ποικίλλει, αλλά και ο αριθμός των ματιών. Όπως γνωρίζουμε, οι άνθρωποι συνήθως έχουν δύο μάτια, οι αράχνες από την άλλη πλευρά έχουν διαφορετικό αριθμό οφθαλμών, με τα περισσότερα είδη να έχουν 8. Κανονικά οι αράχνες έχουν διαφορετικά μεγέθη των διαφορετικών ζευγών οφθαλμών και τα διαφορετικά μεγέθη έχουν διαφορετικά λειτουργίες. Για παράδειγμα, σε εκτινασσόμενες αράχνες 2 μεγαλύτερα μπροστινά προσανατολισμένα μάτια, δίνουν στην αράχνη εξαιρετική οπτική οξύτητα, η οποία χρησιμοποιείται κυρίως για να στοχεύει τη λεία. 6 μικρότερα μάτια έχουν πολύ φτωχότερη ανάλυση, αλλά βοηθούν την αράχνη να αποφύγει πιθανούς κινδύνους. Δύο φωτογραφίες των ματιών μιας εκτινασσόμενης αράχνης και τα μάτια μιας αράχνης λύκου δείχνονται να επιδεικνύουν τη μεταβλητότητα στις τοπολογίες ματιών των αραχνοειδών.
- Οι τοπολογίες των ματιών των αράχνων
-
 Αράχνη λύκου
Αράχνη λύκου -
 Jumping Spider
Jumping Spider


Ανατομία του οπτικού συστήματος
[επεξεργασία]Εμείς οι άνθρωποι είμαστε οπτικά πλάσματα, επομένως τα μάτια μας αποτελούνται απο πολλά στοιχεία. Σε αυτό το κεφάλαιο γίνεται προσπάθεια να περιγραφούν αυτά τα στοιχεία, δίνοντας έτσι κάποια εικόνα για τις ιδιότητες και τη λειτουργικότητα του ανθρώπινου οράματος.
Εισαγωγή στο μάτι - Κόρη, ίρις και φακός
[επεξεργασία]Οι ακτίνες φωτός εισέρχονται στη δομή των ματιών μέσω του μαύρου ανοίγματος ή της κόρης στο μπροστινό μέρος του ματιού. Η μαύρη εμφάνισή της οφείλεται στο γεγονός ότι το φως απορροφάται πλήρως από τον ιστό στο εσωτερικό του ματιού. Μόνο μέσω αυτής της κόρης, μπορεί το φως να εισέλθει μέσα στο μάτι που σημαίνει ότι η ποσότητα του εισερχόμενου φωτός καθορίζεται αποτελεσματικά από το μέγεθος της κόρης. Ένας κυκλικός μυς που ονομάζεται σφιγκτήρας κόρης βρισκεται σε ενα στρωμα που περιβάλλει την κόρη συστέλεται για να σταματά το άνοιγμα του ματιού. Ένας χρωματισμένος σφιγκτήρας που περιβάλλει την κόρη λειτουργεί για να σταματά το άνοιγμα του ματιού. Είναι η ποσότητα της χρωστικής ουσίας σε αυτή την ίριδα, που καθορίζουν τα διάφορα χρώματα των ματιών που βρίσκονται στους ανθρώπους.
Εκτός από αυτό το στρώμα χρωστικής ουσίας, η ίριδα έχει 2 στρώματα ακτινωτών μυών. Ένας κυκλικός μυς που ονομάζεται σφιγκτήρας κόρης σε ένα στρώμα, συμβάλλει ώστε η κόρη να συστέλλεται. Το άλλο στρώμα έχει έναν λείο μυ, που ονομάζεται διαστολέας της κόρης, που συστέλλεται για τη διαστολή αυτής. Ο συνδυασμός αυτών των μυών μπορεί έτσι να διαστέλλει / συστέλλει την κόρη ανάλογα με τις απαιτήσεις ή τις συνθήκες του ατόμου. Οι ακτινωτοί μύες ελέγχονται από ακτινωτές ζόνες, ίνες που επίσης αλλάζουν το σχήμα του φακού και τον κρατούν στη θέση του.
Ο φακός βρίσκεται ακριβώς πίσω από την κόρη. Το σχήμα και τα χαρακτηριστικά του αποκαλύπτουν παρόμοιο σκοπό με εκείνο των φακών της κάμερας, αλλά λειτουργούν με ελαφρώς διαφορετικούς τρόπους. Το σχήμα του φακού ρυθμίζεται από την έλξη των ακτινωτών ζωνών, οι οποίες κατά συνέπεια αλλάζουν το εστιακό μήκος. Μαζί με τον κερατοειδή χιτώνα, ο φακός μπορεί να αλλάξει την εστίαση, πράγμα που καθιστά πράγματι πολύ σημαντική δομή, ωστόσο μόνο το ένα τρίτο της συνολικής οπτικής ισχύος του οφθαλμού οφείλεται στον ίδιο τον φακό. Είναι επίσης το κύριο φίλτρο του ματιού. Οι ίνες φακών αποτελούν το μεγαλύτερο μέρος του υλικού για το φακό, τα οποία είναι μακρά και λεπτά κύτταρα κενά από το μεγαλύτερο μέρος του κυτταρικού μηχανισμού για την προώθηση της διαφάνειας. Μαζί με τις υδατοδιαλυτές πρωτεΐνες που ονομάζονται κρυσταλλίνες, αυξάνουν τον δείκτη διάθλασης του φακού. Οι ίνες διαμορφώνουν τη δομή και το σχήμα του ίδιου του φακού.

Σχηματισμός ακτίνας στο μάτι - Κερατοειδής και προστατευτικός παράγοντας – Σκληρός χιτώνας
[επεξεργασία]
Ο κερατοειδής, υπεύθυνος για τα υπόλοιπα 2/3 της συνολικής οπτικής ισχύος του ματιού, καλύπτει την ίριδα, την κόρη και τον φακό. Εστιάζει τις ακτίνες που περνούν από την ίριδα πριν περάσουν από τον φακό. Ο κερατοειδής χιτώνας έχει πάχος μόνο 0,5 mm και αποτελείται από 5 στρώσεις:
- Επιθήλιο: Ένα στρώμα επιθηλιακού ιστού που καλύπτει την επιφάνεια του
κερατοειδούς.
- Μεμβράνη του Bowman: Ένα παχύ προστατευτικό στρώμα που αποτελείται από
ισχυρές ίνες κολλαγόνου, που διατηρούν το συνολικό σχήμα του κερατοειδούς χιτώνα.
- Stroma: Ένα στρώμα που αποτελείται από παράλληλα ινίδια κολλαγόνου. Αυτό
το στρώμα αποτελεί το 90% του πάχους του κερατοειδούς.
- Η μεμβράνη και το ενδοθήλιο του Descemet: Είναι δύο στρώματα
προσαρμοσμένα στον πρόσθιο θάλαμο του ματιού γεμάτα με υγρό υδατικού υγρού που παράγεται από το ακτινωτό σώμα. Αυτό το υγρό ενυδατώνει τον φακό, καθαρίζει και διατηρεί την πίεση στη σφαίρα του ματιού. Ο θάλαμος, τοποθετημένος μεταξύ του κερατοειδούς και της ίριδας, περιέχει ένα δοκιδωτό σώμα πλέγματος μέσω του οποίου το υγρό αποστραγγίζεται έξω από το κανάλι Schlemm, μέσω του οπίσθιου θαλάμου.
Η επιφάνεια του κερατοειδούς βρίσκεται κάτω από δύο προστατευτικές μεμβράνες, που ονομάζονται σκληρός και η κάψουλα του Tenon. Και τα δύο αυτά προστατευτικά στρώματα καλύπτουν πλήρως το βολβό του ματιού. Ο σκληρός χιτώνας είναι κατασκευασμένος από κολλαγόνο και ελαστικές ίνες, οι οποίες προστατεύουν το μάτι από εξωτερικές βλάβες, και αυτό το στρώμα δημιουργεί και το λευκό του ματιού. Διαπερνάται από νεύρα και αγγεία με τη μεγαλύτερη τρύπα που προορίζεται για το οπτικό νεύρο. Επιπλέον, καλύπτεται από τον επιπεφυκότα, ο οποίος είναι μια σαφής βλεννογόνος μεμβράνη στην επιφάνεια του βολβού. Αυτή η μεμβράνη επίσης ευθυγραμμίζει το εσωτερικό του βλεφάρου. Λειτουργεί ως λιπαντικό και, μαζί με τον δακρυϊκό αδένα, παράγει δάκρυα, που λιπαίνουν και προστατεύουν το μάτι. Το υπόλοιπο προστατευτικό στρώμα, το βλέφαρο, λειτουργεί επίσης για να διαχέει αυτό το λιπαντικό γύρω.
Μετακίνηση των οφθαλμών - εξοφθαλμικοί μύες
[επεξεργασία]Ο βολβός μετακινείται από μια πολύπλοκη μυϊκή δομή των εξοφθαλμικών μυών που αποτελείται από τέσσερις μύες του ορθού - κατώτερου, μέσου, πλευρικού και ανώτερου και δύο λοξού - κατώτερου και ανώτερου. Η τοποθέτηση αυτών των μυών παρουσιάζεται παρακάτω, μαζί με τις λειτουργίες τους:

Όπως βλέπουμε, οι εξω-οφθαλμικοί μύες (2,3,4,5,6,8) συνδέονται με τον
σκληρό χιτώνα του ματιού και προέρχονται από τον δακτύλιο του Zinn, έναν
ινώδη τένοντα που περιβάλλει το οπτικό νεύρο. Ένα σύστημα τροχαλίας
δημιουργείται με το trochlea να λειτουργεί ως τροχαλία και τον ανώτερο λοξό
μυ όπως το σχοινί, αυτό είναι απαραίτητο για να ανακατευθύνει τη μυϊκή
δύναμη με τον σωστό τρόπο. Οι υπόλοιποι εξοφθαλμικοί μύες έχουν άμεση οδό
προς τα μάτια και επομένως δεν σχηματίζουν αυτά τα συστήματα τροχαλιών.
Χρησιμοποιώντας αυτούς τους εξοφθαλμικούς μύες, το μάτι μπορεί να
περιστρέφεται προς τα επάνω, προς τα κάτω, προς τα αριστερά, προς τα δεξιά
και εναλλακτικές κινήσεις είναι δυνατές ως συνδυασμός αυτών.
Άλλες κινήσεις είναι επίσης πολύ σημαντικές για να μπορέσουμε να δούμε. Οι κινήσεις βροχής επιτρέπουν τη σωστή λειτουργία της διοφθαλμικής όρασης. Οι ασυνείδητες ταχείς κινήσεις που ονομάζονται σακκάδες είναι απαραίτητες για να μπορούν οι άνθρωποι να εστιάζονται σε ένα αντικείμενο. Η σακκάδα είναι ένα είδος θορυβώδους κίνησης που εκτελείται όταν τα μάτια σαρώνουν το οπτικό πεδίο, προκειμένου να μετατοπίσουν ελαφρά το σημείο εστίασης. Όταν ακολουθείτε ένα κινούμενο αντικείμενο με το βλέμμα σας, τα μάτια σας εκτελούν αυτό που αναφέρεται ως ομαλή αναζήτηση. Πρόσθετες ακούσιες κινήσεις που ονομάζονται νυσταγμός προκαλούνται από σήματα από το αιθουσαίο σύστημα, μαζί σχηματίζουν τα αιθουσα-οφθαλμικά αντανακλαστικά. Το στέλεχος του εγκεφάλου ελέγχει όλες τις κινήσεις των ματιών, με διαφορετικές περιοχές υπεύθυνες για διαφορετικές κινήσεις.
- Pons: Ταχείες οριζόντιες κινήσεις, όπως σακκάδες ή νυσταγμός.
- Mesencephalon: Κάθετες και στρεπτικές κινήσεις.
- Υποσχηματισμός: Εξομάλυνση
- Ο πυρήνας Edinger-Westphal: Οι κινήσεις της βροχής.
Το σημείο που δημιουργείται η όραση - Ο αμφιβληστροειδής
[επεξεργασία]
Πριν από τη μετάδοσή του, το εισερχόμενο ΗΜ περνά μέσα από τον κερατοειδή χιτώνα, τον φακό και την κηλίδα. Αυτές οι δομές λειτουργούν επίσης ως φίλτρα για τη μείωση των ανεπιθύμητων EM, προστατεύοντας έτσι το μάτι από τη βλαβερή ακτινοβολία. Η απόκριση φιλτραρίσματος καθενός από αυτά τα στοιχεία μπορεί να φανεί στο σχήμα "Φιλτράρισμα του φωτός που εκτελείται από τον κερατοειδή χιτώνα, το φακό και το χρωστικό επιθήλιο". Όπως μπορεί κανείς να παρατηρήσει, ο κερατοειδής εξασθενεί τα χαμηλότερα μήκη κύματος, αφήνοντας τα υψηλότερα μήκη κύματος σχεδόν ανέγγιχτα. Ο φακός μπλοκάρει περίπου το 25% του EM κάτω από 400nm και περισσότερο από 50% κάτω από 430nm. Τέλος, το τελικό στάδιο φιλτραρίσματος πριν από τη λήψη φωτογραφιών επηρεάζει περίπου το 30% του ΗΜ μεταξύ 430nm και 500nm.
Ένα μέρος του ματιού, το οποίο σηματοδοτεί τη μετάβαση από μη ευαίσθητη περιοχή σε φωτοευαίσθητη περιοχή, ονομάζεται ora serrata. Η φωτοευαίσθητη περιοχή αναφέρεται ως αμφιβληστροειδής, η οποία είναι η αισθητηριακή δομή στο πίσω μέρος του ματιού. Ο αμφιβληστροειδής αποτελείται από πολλαπλά στρώματα που παρουσιάζονται παρακάτω με εκατομμύρια φωτοϋποδοχείς που ονομάζονται ραβδία και κώνοι, οι οποίοι συλλαμβάνουν τις ακτίνες φωτός και τις μετατρέπουν σε ηλεκτρικές παλμούς. Η μετάδοση αυτών των παρορμήσεων ξεκινάει νευρικά από τα γαγγλιακά κύτταρα και διέρχεται μέσω του οπτικού νεύρου, της μοναδικής οδού μέσω της οποίας οι πληροφορίες φεύγουν από το μάτι.
Δομή του αμφιβληστροειδούς που περιλαμβάνει τα κυριότερα κυτταρικά συστατικά: RPE: επιθήλιο χρωστικής αμφιβληστροειδούς, OS: εξωτερικό τμήμα των κυττάρων φωτοϋποδοχέα. IS: εσωτερικό τμήμα των κυττάρων φωτοϋποδοχέα. ONL: εξωτερική πυρηνική στρώση. OPL: εξωτερικό πλεγματοειδές στρώμα. INL: εσωτερική πυρηνική στιβάδα IPL: εσωτερική πλευρική στρώση. GC: στοιβάδα κυττάρων γαγγλίου, Ρ: κύτταρο επιθηλίου χρωστικής, ΒΜ: Bruch-Membran; R: ραβδία. Γ: κώνοι. H: οριζόντιο κύτταρο; Β: διπολικό κύτταρο. Μ: κύτταρο Müller; Α: κύτταρο αμακρίνης. G: κύτταρο γαγγλίων. AX: Axon; βέλος: Μεμβρανικά όρια εξωτερικά.

Μια εννοιολογική απεικόνιση της δομής του αμφιβληστροειδούς φαίνεται στα δεξιά. Όπως μπορούμε να δούμε, υπάρχουν πέντε κύριοι τύποι κυττάρων:
- κύτταρα φωτοϋποδοχέα
- οριζόντια κύτταρα
- διπολικά κύτταρα
- κύτταρα αμετρίνης
- κύτταρα γαγγλίων
Τα κύτταρα φωτοϋποδοχέα μπορούν να υποδιαιρεθούν περαιτέρω σε δύο κύριους τύπους που ονομάζονται ραβδία και κώνοι. Οι κώνοι είναι πολύ λιγότεροι από τις ραβδίους στα περισσότερα μέρη του αμφιβληστροειδούς, αλλά υπάρχει μια τεράστια συσσώρευση αυτών στην ωχρά κηλίδα, ειδικά στο κεντρικό τμήμα που ονομάζεται φούντα. Σε αυτή την κεντρική περιοχή, κάθε φωτοευαίσθητος κώνος συνδέεται με ένα γαγγλιοκύτταρο. Επιπλέον, οι κώνοι σε αυτήν την περιοχή είναι ελαφρώς μικρότεροι από τον μέσο όρο του κώνου, πράγμα που σημαίνει ότι υπάρχουν περισσότεροι κώνους ανά περιοχή. Λόγω αυτού του λόγου και της υψηλής πυκνότητας των κώνων, αυτό είναι το σημείο με την υψηλότερη οπτική οξύτητα.
Υπάρχουν 3 τύποι ανθρώπινων κώνων, έκαστος των κώνων που ανταποκρίνονται σε συγκεκριμένο εύρος μηκών κύματος, εξαιτίας τριών τύπων χρωστικής που ονομάζεται φωτοψίνη. Κάθε χρωστική ουσία είναι ευαίσθητη στο κόκκινο, μπλε ή πράσινο μήκος κύματος του φωτός, έτσι έχουμε μπλε, πράσινους και κόκκινους κώνους, που ονομάζονται επίσης S-, M- και L-cones για την ευαισθησία τους σε βραχύ, μεσαίο και μακρύ μήκος κύματος αντίστοιχα. Αποτελείται από πρωτεΐνη που ονομάζεται opsin και δεσμευμένο χρωμόρφο ονομάζεται αμφιβληστροειδής. Τα κυριότερα δομικά στοιχεία του κυττάρου του κώνου είναι το συναπτικό τερματικό, το εσωτερικό και το εξωτερικό τμήμα, ο εσωτερικός πυρήνας και τα μιτοχόνδρια.
Οι φασματικές ευαισθησίες των 3 τύπων κώνων:
• 1. Οι κώνοι S απορροφούν το φως βραχέων κυμάτων, δηλαδή το μπλε-ιώδες φως. Το μέγιστο μήκος κύματος απορρόφησης για τους S-κώνους είναι 420 nm • 2. Οι κώνοι M απορροφούν μπλε-πράσινο έως κίτρινο φως. Στην περίπτωση αυτή το μέγιστο μήκος κύματος απορρόφησης είναι 535nm • 3. Οι L-κώνες απορροφούν το κίτρινο έως το κόκκινο φως. Το μέγιστο μήκος κύματος απορρόφησης είναι 565nm

Το εσωτερικό τμήμα περιέχει οργανίδια και τον πυρήνα και τα οργανίδια του κυττάρου. Η χρωστική βρίσκεται στο εξωτερικό τμήμα, προσαρτημένη στη μεμβράνη ως πρωτεΐνες διαμεμβράνης εντός των εισαγωγών της κυτταρικής μεμβράνης που σχηματίζουν τους μεμβρανώδεις δίσκους, οι οποίοι είναι σαφώς ορατοί στο σχήμα που απεικονίζει τη βασική δομή των κυττάρων ραβδίου και κώνου. Οι δίσκοι μεγιστοποιούν την περιοχή λήψης των κυψελών.
Οι φωτοϋποδοχείς κώνων πολλών σπονδυλωτών περιέχουν σφαιρικά οργανίδια που ονομάζονται σταγονίδια τα οποία πιστεύεται ότι αποτελούν ενδοφθάλμια φίλτρα που μπορούν να χρησιμεύσουν για την αύξηση της αντίθεσης, τη μείωση της απόφραξης και τη μείωση των χρωματικών παρεκκλίσεων που προκαλούνται από τη μεταβολή του μιτοχονδριακού μεγέθους από την περιφέρεια στα κέντρα.
Οι ραβδία έχουν δομή παρόμοια με τους κώνους, ωστόσο περιέχουν την χρωστική ροδόψιν, η οποία τους επιτρέπει να ανιχνεύουν φως χαμηλής έντασης και τις καθιστά 100 φορές πιο ευαίσθητες από τους κώνους. Η ροδοψίνη είναι η μόνη χρωστική ουσία που βρίσκεται σε ανθρώπινες ραβδίους και βρίσκεται στην εξωτερική πλευρά του χρωστικού επιθηλίου, το οποίο παρόμοια με τους κώνους μεγιστοποιεί την περιοχή απορρόφησης χρησιμοποιώντας μια δομή δίσκου. Ομοίως με τους κώνους, ο συνοπτικός άκρων του κυττάρου συνδέεται με ένα διπολικό κύτταρο και τα εσωτερικά και εξωτερικά τμήματα συνδέονται με το cilium.
Η χρωστική ροδοψίνη απορροφά το φως μεταξύ 400-600nm, με μέγιστη απορρόφηση γύρω στα 500nm. Αυτό το μήκος κύματος αντιστοιχεί στο πράσινο-μπλε φως που σημαίνει ότι τα μπλε χρώματα εμφανίζονται πιο έντονα σε σχέση με τα κόκκινα χρώματα τη νύχτα.

Τα EM κύματα με μήκη κύματος εκτός της περιοχής των 400 - 700 nm δεν ανιχνεύονται ούτε από ραβδίους ούτε από κώνους, πράγμα που τελικά σημαίνει ότι δεν είναι ορατά για τον άνθρωπο. Τα οριζόντια κύτταρα καταλαμβάνουν το εσωτερικό πυρηνικό στρώμα του αμφιβληστροειδούς. Υπάρχουν δύο τύποι οριζόντιων κυψελών και οι δύο τύποι υπερπολικοί σε απόκριση στο φως δηλ. Γίνονται πιο αρνητικοί. Ο τύπος Α αποτελείται από έναν υπότυπο που ονομάζεται HII-H2 ο οποίος αλληλεπιδρά με κυρίους S-κώνους. Τα κύτταρα τύπου Β έχουν έναν υπότυπο που ονομάζεται HIH1, ο οποίος διαθέτει δέντρο δεντρίτη κι ένα άξονα. Οι πρώτες επαφές αποτελούνται κυρίως από κύτταρα Μ και L-κώνου και τα τελευταία κύτταρα ραβδίου. Οι επαφές με κώνους είναι κατασκευασμένες κυρίως από απαγορευτικές συνάψεις, ενώ τα ίδια τα κύτταρα ενώνονται σε ένα δίκτυο με κόμβους διακένου.

Τα διπολικά κύτταρα απλώνουν μονούς δενδρίτες στο εξωτερικό πλεγματοειδές στρώμα και το περικάρυο, τα κυτταρικά σώματα τους, βρίσκονται στο εσωτερικό πυρηνικό στρώμα. Οι δενδρίτες αλληλοσυνδέονται αποκλειστικά με κώνους και ραβδίους και διαφοροποιούνται μεταξύ ενός διπολικού κυττάρου και εννέα ή δέκα κώνων διπολικών κυττάρων. Αυτά τα κύτταρα διακλαδίζονται με κύτταρα αμακρίνης ή γαγγλίων στην εσωτερική πλεγματοειδή στιβάδα με τη χρήση ενός αξόνου. Τα διπολικά κύτταρα της ραβδίου συνδέονται με συσπάσεις τριάδας ή με κυψελίδες 18-70. Οι άξονες τους εξαπλώνονται γύρω από τα εσωτερικά πλεγματοειδή στρώματα συναπτικών ακροδεκτών, τα οποία περιέχουν συνάψεις κορδέλας και έρχονται σε επαφή με ένα ζεύγος κυτταρικών διεργασιών σε δυαδικές συνάψεις. Συνδέονται με κύτταρα γαγγλίων με τις κυτταρικές συνδέσεις της αμεκρίνης.
Τα κύτταρα Amecrine μπορούν να βρεθούν στην εσωτερική πυρηνική στιβάδα και στο στρώμα γαγγλιοκυττάρων του αμφιβληστροειδούς. Περιστασιακά βρίσκονται στο εσωτερικό πλεγματοειδές στρώμα, όπου λειτουργούν ως διαμορφωτές σήματος. Έχουν ταξινομηθεί ως στενό πεδίο, μικρό πεδίο, μεσαίο πεδίο ή ευρύ πεδίο ανάλογα με το μέγεθός τους. Ωστόσο, υπάρχουν πολλές ταξινομήσεις που οδηγούν σε πάνω από 40 διαφορετικούς τύπους αμετρικών κυττάρων.
Τα κύτταρα γαγγλίων είναι οι τελικοί πομποί οπτικού σήματος από τον αμφιβληστροειδή στον εγκέφαλο. Τα πιο συνηθισμένα κύτταρα γαγγλίων στον αμφιβληστροειδή είναι το κύτταρο των γαγγλίων και το γαγγλιακό κύτταρο ομπρέλας. Το σήμα μετά τη διέλευση από όλες τις στιβάδες αμφιβληστροειδούς μεταφέρεται σε αυτά τα κύτταρα τα οποία είναι το τελικό στάδιο της αλυσίδας επεξεργασίας του αμφιβληστροειδούς. Όλες οι πληροφορίες συλλέγονται εδώ προωθούμενες στις αμφιβληστροειδικές νευρικές ίνες και στα οπτικά νεύρα. Το σημείο όπου οι άξονες των γαγγλίων συγχωνεύονται για να δημιουργήσουν οπτικό νεύρο ονομάζεται οπτικός δίσκος. Αυτό το νεύρο κατασκευάζεται κυρίως από τους νευραξονικούς νευράξονες του αμφιβληστροειδούς και τα κύτταρα Portort. Η πλειονότητα των αξόνων μεταδίδει δεδομένα στον πλευρικό γονιδιακό πυρήνα, ο οποίος είναι ένας σύνδεσμος τερματισμού για τα περισσότερα τμήματα του νεύρου και ο οποίος προωθεί την πληροφορία στον οπτικό φλοιό. Μερικά γαγγλιακά κύτταρα αντιδρούν επίσης στο φως, αλλά επειδή αυτή η απόκριση είναι πιο αργή από αυτή των ράβδων και των κώνων, πιστεύεται ότι σχετίζεται με την ανίχνευση των επιπέδων φωτός του περιβάλλοντος και την προσαρμογή του βιολογικού ρολογιού.
Επεξεργασία σήματος
[επεξεργασία]Όπως αναφέρθηκε προηγουμένως ο αμφιβληστροειδής είναι το κύριο στοιχείο στο μάτι, επειδή περιέχει όλα τα φωτοευαίσθητα κύτταρα. Χωρίς αυτό, το μάτι θα ήταν συγκρίσιμο με μια ψηφιακή κάμερα χωρίς τον αισθητήρα CCD (Charge Coupled Device). Αυτό το μέρος εξηγεί πώς ο αμφιβληστροειδής αντιλαμβάνεται το φως, πώς μεταδίδεται το οπτικό σήμα στον εγκέφαλο και πώς ο εγκέφαλος επεξεργάζεται το σήμα για να σχηματίσει αρκετές πληροφορίες για τη λήψη αποφάσεων.
Δημιουργία των αρχικών σημάτων - Λειτουργία φωτοαισθητήρα
[επεξεργασία]Η όραση γεννάται πάντα με το φως να χτυπά τα φωτοευαίσθητα κύτταρα που βρίσκονται στον αμφιβληστροειδή χιτώνα. Οι οπτικές χρωστικές που απορροφούν το φως, μια ποικιλία ενζύμων και πομπών σε ραβδίους αμφιβληστροειδούς και κώνοι θα ξεκινήσουν τη μετατροπή από ορατά ΗΜ ερεθίσματα σε ηλεκτρικούς παλμούς, με μια διαδικασία γνωστή ως φωτοηλεκτρική μεταγωγή.
Χρησιμοποιώντας ραβδία ως παράδειγμα, τα εισερχόμενα ορατά ΗΜ συναντούν μόρια ροδοψίνης, διαμεμβρανικά μόρια που βρίσκονται στη δομή του εξωτερικού δίσκου των ράβδων. Κάθε μόριο ροδοψίνης αποτελείται από ένα σύμπλεγμα ελίκων που ονομάζεται opsin που περιβάλλει και περιβάλλει τον 11-cis αμφιβληστροειδή, το οποίο είναι το τμήμα του μορίου που θα αλλάξει λόγω της ενέργειας από τα εισερχόμενα φωτόνια. Σε βιολογικά μόρια, τμήματα ή τμήματα μορίων που θα προκαλέσουν αλλαγές διαμόρφωσης λόγω αυτής της ενέργειας αναφέρονται μερικές φορές ως χρωμοφόρα. Ο 11-cis αμφιβληστροειδής ισορροπεί σε απόκριση στην εισερχόμενη ενέργεια, μετατρέποντας τον αμφιβληστροειδή (all-trans αμφιβληστροειδής), η οποία ωθεί τις έλικες opsin πιο μακριά, προκαλώντας την αποκάλυψη ειδικών αντιδραστικών θέσεων.
Αυτό το "ενεργοποιημένο" μόριο ροδοψίνης αναφέρεται μερικές φορές ως Μεθαροδοψίνη II. Από αυτό το σημείο, ακόμα και αν σταματήσει η διέγερση του ορατού φωτός, η αντίδραση θα συνεχιστεί. Η μεθαροδοψίνη II μπορεί στη συνέχεια να αντιδρά με περίπου 100 μόρια μιας πρωτεΐνης Gs που ονομάζεται μεταγωγή, η οποία στη συνέχεια οδηγεί σε α και β; μετά την μετατροπή του ΑΕΠ σε GTP. Το ενεργοποιημένο ως-ΟΤΡ στη συνέχεια δεσμεύεται με cGMP-φωσφοδιεστεράση (PDE), καταστέλλοντας τις κανονικές λειτουργίες ανταλλαγής ιόντων, με αποτέλεσμα τη χαμηλή συγκέντρωση ιόντων κατιόντων κυτοσόλης και επομένως μια αλλαγή στην πόλωση του κυττάρου.
Η φυσική αντίδραση φωτοηλεκτρικής μεταγωγής έχει μια καταπληκτική δύναμη ενίσχυσης. Ένα μόριο αμφιβληστροειδούς ροδοψίνης που ενεργοποιείται από ένα μοναδικό κβαντικό φως προκαλεί την υδρόλυση μέχρι 106 μορίων cGMP ανά δευτερόλεπτο.
Μεταγωγή φωτογραφιών
[επεξεργασία]- Ένα φωτεινό φωτόνιο αλληλεπιδρά με τον αμφιβληστροειδή σε ένα φωτοδέκτη. Ο αμφιβληστροειδής υποβάλλεται σε ισομερισμό, αλλάζοντας από τη διαμόρφωση από 11-cis σε all-trans.
- Ο αμφιβληστροειδής δεν ταιριάζει πλέον στη θέση δέσμευσης opsin.
- Επομένως, το Opsin υφίσταται μεταβολική διαμόρφωση σε μεταροδoψίνη II.
- Η μεταορδοψίνη II είναι ασταθής και διασπάται, αποδίδοντας οψίνη και all-trans αμφιβληστροειδούς.
- Η οψίνη ενεργοποιεί τη ρυθμιστική πρωτεΐνη transducin. Αυτό προκαλεί την αποικοδόμηση της transducin από το δεσμευμένο ΑΕΠ της και τη δέσμευση του GTP, τότε η άλφα υπομονάδα της transducin διαχωρίζεται από τις βήτα και γάμμα υπομονάδες, ενώ η GTP εξακολουθεί να συνδέεται με την άλφα υπομονάδα.
- Το σύμπλεγμα άλφα υπομονάδας-GTP ενεργοποιεί τη φωσφοδιεστεράση.
- Η φωσφοδιεστεράση διασπά το cGMP σε 5'-GMP. Αυτό μειώνει τη συγκέντρωση της cGMP και κατά συνέπεια τα κανάλια νατρίου κλείνουν.
- Το κλείσιμο των διαύλων νατρίου προκαλεί υπερπόλωση του κυττάρου λόγω του συνεχούς ρεύματος καλίου.
- Η υπερπόλωση του κυττάρου προκαλεί το κλείσιμο των τασεοελεγχόμενων διαύλων ασβεστίου.
- Καθώς πέφτει το επίπεδο ασβεστίου στο κύτταρο φωτοϋποδοχέα, μειώνεται επίσης η ποσότητα του γλουταμικού νευροδιαβιβαστή που απελευθερώνεται από το κύτταρο. Αυτό συμβαίνει επειδή το ασβέστιο απαιτείται για τα κυστίδια που περιέχουν γλουταμινικό να συγχωνευθούν με την κυτταρική μεμβράνη και να απελευθερώσουν τα περιεχόμενά τους.
- Μία μείωση της ποσότητας γλουταμινικού που απελευθερώνεται από τους φωτοϋποδοχείς προκαλεί αποπόλωση των κεντρικών διπολικών κυττάρων (ραβδίου και κώνου στα διπολικά κύτταρα) και υπερπόλωση κώνου εκτός διπολικών κυττάρων.
Χωρίς ορατή ΗΜ διέγερση, τα κύτταρα ραβδίου που περιέχουν ένα κοκτέιλ ιόντων, πρωτεϊνών και άλλων μορίων, έχουν διαφορές δυναμικού μεμβράνης περίπου -40mV. Σε σύγκριση με άλλα νευρικά κύτταρα, αυτό είναι αρκετά υψηλό (-65mV). Σε αυτή την κατάσταση, ο γλουταμικός νευροδιαβιβαστής απελευθερώνεται συνεχώς από τα τερματικά του νευραξονίου και απορροφάται από τα γειτονικά διπολικά κύτταρα. Με την εισερχόμενη ορατή ΗM και την προαναφερθείσα καταρράκτη, η διαφορά δυναμικού πέφτει στα -70mV. Αυτός ο υπερπολισμός του κυττάρου προκαλεί μείωση της ποσότητας του απελευθερούμενου γλουταμικού, επηρεάζοντας έτσι τη δραστηριότητα των διπολικών κυττάρων και στη συνέχεια τα ακόλουθα στάδια στην οπτική οδό. Παρόμοιες διεργασίες υπάρχουν στα κωνικά-κύτταρα και στα φωτοευαίσθητα γαγγλιακά κύτταρα, αλλά κάνουν χρήση διαφορετικών οψινών. Η φωτοψίνη Ι έως ΙΙΙ (κιτρινοπράσινη, πράσινη και μπλε-βιολετί αντίστοιχα) βρίσκεται στα τρία διαφορετικά κωνικά κύτταρα και η μελανοψίνη (μπλε) μπορεί να βρεθεί στα φωτοευαίσθητα γαγγλιακά κύτταρα.
Επεξεργασία σημάτων στον αμφιβληστροειδή
[επεξεργασία]Διαφορετικά διπολικά κύτταρα αντιδρούν διαφορετικά στις αλλαγές στο απελευθερούμενο γλουταμικό. Τα λεγόμενα διπολικά κύτταρα ON και OFF χρησιμοποιούνται για να σχηματίσουν την άμεση ροή σήματος από κώνους σε διπολικά κύτταρα. Τα διπολικά κύτταρα ON θα αποπολωθούν με ορατή ΗΜ διέγερση και θα ενεργοποιηθούν τα αντίστοιχα ΟΝ γαγγλιοκύτταρα. Από την άλλη πλευρά, τα διπολικά κύτταρα OFF είναι υπερπολικά πολωμένα με την ορατή ΗM διέγερση, και τα κύτταρα γαγγλίων OFF αναστέλλονται. Αυτή είναι η βασική οδός της απευθείας ροής σήματος. Η πλευρική ροή σήματος θα ξεκινήσει από τα ραβδία και στη συνέχεια θα μεταβεί στα διπολικά κύτταρα, τα κύτταρα της αμακρίνης και τα διπολικά κύτταρα OFF που αναστέλλονται από τα κύτταρα Rodamacrine και τα διπολικά κύτταρα ON θα διεγερθούν μέσω μιας ηλεκτρικής συνάψεως, μετά από όλα τα προηγούμενα στα βήματα, το σήμα θα φτάσει στα κύτταρα γαγγλίων ON ή OFF και θα καθοριστεί ολόκληρη η πορεία της ροής σήματος.
Όταν το δυναμικό δράσης (ΔΔ) είναι στο ON, τα κύτταρα των γαγγλίων θα ενεργοποιηθούν από το ορατό ΗΜ ερέθισμα. Η συχνότητα ΔΔ θα αυξηθεί όταν αυξηθεί το δυναμικό του αισθητήρα. Με άλλα λόγια, το ΔΔ εξαρτάται από το εύρος του δυναμικού του αισθητήρα. Η περιοχή των γαγγλιοκυττάρων όπου τα διεγερτικά και ανασταλτικά αποτελέσματα επηρεάζουν τη συχνότητα ΔΔ ονομάζεται πεδίο δεκτικότητας (ΠΔ). Γύρω από τα κύτταρα των γαγγλίων, το ΠΔ συνήθως αποτελείται από δύο περιοχές: την κεντρική ζώνη και την περιφερειακή ζώνη που μοιάζει με δακτύλιο. Διακρίνονται κατά τη διάρκεια της ορατής ΗΜ προσαρμογής. Μια ορατή ΗM διέγερση στην κεντρική ζώνη θα μπορούσε να οδηγήσει σε αύξηση της συχνότητας ΔΔ και η διέγερση στη ζώνη της περιφέρειας θα μειώσει τη συχνότητα ΔΔ. Όταν η πηγή φωτός είναι απενεργοποιημένη, εμφανίζεται η διέγερση. Έτσι, το όνομα του πεδίου ON (κεντρικό πεδίο ON) αναφέρεται σε αυτό το είδος περιοχής. Φυσικά, το ΠΔ των κυττάρων των γαγγλίων OFF ενεργεί με τον αντίθετο τρόπο και επομένως ονομάζεται "πεδίο OFF" (κεντρικό πεδίο απενεργοποιημένο). Τα ΠΔ οργανώνονται από τα οριζόντια κελιά. Η ώθηση στην περιοχή της περιφέρειας θα παρορμηθεί και θα μεταδοθεί στην κεντρική περιοχή και εκεί θα σχηματιστεί η αποκαλούμενη αντίθεση ερεθίσματος. Αυτή η λειτουργία θα κάνει το σκοτάδι να φαίνεται πιο σκοτεινό και το φως πιο φωτεινό. Εάν ολόκληρο το ΠΔ εκτίθεται στο φως, η ώθηση της κεντρικής περιοχής θα κυριαρχήσει.
Μετάδοση σήματος στο φλοιό
[επεξεργασία]Όπως αναφέρθηκε προηγουμένως, οι άξονες των κυττάρων των γαγγλιοκυττάρων συγκλίνουν στον οπτικό δίσκο του αμφιβληστροειδούς, σχηματίζοντας το οπτικό νεύρο. Αυτές οι ίνες τοποθετούνται μέσα στη δέσμη σε συγκεκριμένη σειρά. Οι ίνες από τη ζώνη της ωχράς κηλίδας του αμφιβληστροειδούς είναι στο κεντρικό τμήμα και εκείνες από το προσωρινό ήμισυ του αμφιβληστροειδούς καταλαμβάνουν το περιφερικό τμήμα. Παρουσιάζεται μερική αποκοπή ή διασταύρωση όταν αυτές οι ίνες είναι έξω από την κοιλότητα των ματιών. Οι ίνες από τα ρινικά μισά του κάθε αμφιβληστροειδούς περνούν στα αντίθετα μισά και εκτείνονται στον εγκέφαλο. Εκείνοι από τα αντίθετα μισά παραμένουν χωρίς περικοπή. Αυτή η μερική διασταύρωση ονομάζεται οπτικό σχίσμα και τα οπτικά νεύρα πέρα από αυτό το σημείο ονομάζονται οπτικοί οδοί, κυρίως για να τα διακρίνουν από τα νεύρα ενός αμφιβληστροειδούς. Η λειτουργία της μερικής διασταύρωσης είναι η μετάδοση του δεξιού οπτικού πεδίου που παράγεται από τα δύο μάτια μόνο στο αριστερό ήμισυ του εγκεφάλου και αντιστρόφως. Επομένως, οι πληροφορίες από το δεξιό ήμισυ του σώματος και το σωστό οπτικό πεδίο μεταδίδονται στο αριστερό μέρος του εγκεφάλου όταν φθάνουν στο οπίσθιο τμήμα του προ-εγκεφάλου (διεγκεφάλου).

Ο αναμεταδότης πληροφοριών μεταξύ των ινών των οπτικών οδών και των
νευρικών κυττάρων εμφανίζεται στα πλευρικά όργανα γονιδιώματος, το κεντρικό
τμήμα της επεξεργασίας οπτικού σήματος, που βρίσκεται στον θάλαμο του
εγκεφάλου. Από εδώ η πληροφορία μεταφέρεται στα νευρικά κύτταρα στον ινιακό
φλοιό της αντίστοιχης πλευράς του εγκεφάλου. Οι συνδέσεις από τον
αμφιβληστροειδή προς τον εγκέφαλο μπορούν να διαχωριστούν σε μια
«παραβοκυτταρική οδό» και μια «μαγνητική κυτταρική οδό». Τα μονοκύτταρα
μονοπάτια σηματοδοτούν χρώμα και λεπτή λεπτομέρεια, ενώ τα μαγνητικά
μονοπάτια ανιχνεύουν ταχέως κινούμενα ερεθίσματα.

Τα σήματα από τις τυποποιημένες ψηφιακές κάμερες αντιστοιχούν περίπου σε εκείνα της μονοκυτταρικής οδού. Για να προσομοιώσουν τις αποκρίσεις των μονοκυτταρικών μονοπατιών, οι ερευνητές αναπτύσσουν νευρομορφικά αισθητήρια συστήματα, τα οποία προσπαθούν να μιμηθούν υπολογισμό με βάση ακίδα σε νευρικά συστήματα. Χρησιμοποιούν έτσι ένα σχήμα που ονομάζεται "ΑddressΕvent representation" για τη μετάδοση σήματος στα νευρομορφικά ηλεκτρονικά συστήματα (Liu και Delbruck 2010 [1]).
Ανατομικά, τα κύτταρα γαγγλίων Magno και Parvo αμφιβληστροειδούς προβάλλουν αντιστοίχως σε 2 κοιλιακά μαγνητικά στρώματα και 4 ραχιαία μεσοκυττάρια στρώματα του Lateral Geniculate Nucleus (LGN). Κάθε ένα από τα έξι στρώματα LGN λαμβάνει εισροές είτε από το ομόπλευρο είτε από το αντίθετο μάτι, δηλαδή τα γαγγλιακά κύτταρα του αριστερού οφθαλμού περνούν και προβάλλουν στο στρώμα 1, 4 και 6 του δεξιού LGN και το έργο των κυττάρων των δεξιών γαγγλίων (χωρίς περικοπή ) στο στρώμα 2, 3 και 5. Από εκεί διαχωρίζονται οι πληροφορίες από το δεξί και το αριστερό μάτι. Αν και η ανθρώπινη όραση συνδυάζεται με δύο μισά του αμφιβληστροειδούς και το σήμα επεξεργάζεται από τα αντίθετα εγκεφαλικά ημισφαίρια, το οπτικό πεδίο θεωρείται ως ομαλή και πλήρης μονάδα. Ως εκ τούτου οι δύο οπτικές φλοιώδεις περιοχές θεωρούνται ότι είναι στενά συνδεδεμένες. Αυτή η σύνδεση, που ονομάζεται corpus callosum, αποτελείται από νευρώνες, άξονες και δενδρίτες. Επειδή οι δενδρίτες σχηματίζουν συναπτικές συνδέσεις στα σχετικά σημεία των ημισφαιρίων, η ηλεκτρική προσομοίωση κάθε σημείου σε ένα ημισφαίριο δείχνει προσομοίωση του διασυνδεδεμένου σημείου στο άλλο ημισφαίριο. Η μόνη εξαίρεση σε αυτόν τον κανόνα είναι ο πρωταρχικός οπτικός φλοιός.
Οι συνάψεις γίνονται από την οπτική οδό στις αντίστοιχες στρώσεις του πλευρικού γονιδιακού σώματος. Στη συνέχεια, οι άξονες αυτών των νευρικών κυττάρων τρίτης τάξης περνούν μέχρι τη σχισμή της χολινής σε κάθε ινιακό λοβό του εγκεφαλικού φλοιού. Επειδή οι ζώνες των λευκών ινών και των ζέυγη νευραξόνων από τα νευρικά κύτταρα του αμφιβληστροειδούς περνούν μέσα από αυτό, ονομάζεται φλοιός του ραβδωτού σώματος, ο οποίος παρεμπιπτόντως είναι ο πρωταρχικός μας οπτικός φλοιός, μερικές φορές γνωστός ως V1. Σε αυτό το σημείο, οι παρορμήσεις από τα ξεχωριστά μάτια συγκλίνουν σε κοινούς φλοιώδεις νευρώνες, που στη συνέχεια επιτρέπουν την πλήρη εισαγωγή και από τα δύο μάτια σε μια περιοχή για να χρησιμοποιηθούν για την αντίληψη και την κατανόηση. Η αναγνώριση των μοτίβων είναι μια πολύ σημαντική λειτουργία αυτού του συγκεκριμένου μέρους του εγκεφάλου, του οποίου βλάβες μπορύν να προκαλέσουν προβλήματα στην οπτική αναγνώριση ή τύφλωση.
Βάσει του διατεταγμένου τρόπου με τον οποίο οι ίνες της οπτικής οδού περνούν πληροφορίες στα πλευρικά σωμάτια του γονιδιώματος και μετά που περνούν στην περιοχή των ραβδώσεων, αν βρεθεί μία διέγερση ενός σημείου στον αμφιβληστροειδή, η απόκριση που παράγεται ηλεκτρικά και στο αμφίπλευρο σώμα γονιδιώματος και ο πτερυγικός φλοιός θα βρεθεί σε μια μικρή περιοχή στο συγκεκριμένο σημείο του αμφιβληστροειδούς. Αυτός είναι ένας προφανής τρόπος σημειακής επεξεργασίας σημάτων. Και αν διεγερθεί ολόκληρος ο αμφιβληστροειδής, οι αποκρίσεις θα συμβούν και στα δύο πλευρικά όργανα γονιδιώματος και στην περιοχή της φαιάς ύφανσης του σχοινιού. Είναι δυνατό να χαρτογραφηθεί αυτή η περιοχή του εγκεφάλου στα πεδία του αμφιβληστροειδούς, ή συνηθέστερα στα οπτικά πεδία.
Οποιαδήποτε περαιτέρω βήματα σε αυτό το μονοπάτι είναι πέρα από το πεδίο αυτού του βιβλίου. Είναι βέβαιο ότι υπάρχουν πολλά ακόμα επίπεδα και κέντρα, εστιάζοντας σε συγκεκριμένα συγκεκριμένα καθήκοντα, όπως για παράδειγμα χρώμα, προσανατολισμούς, χωρικές συχνότητες, συναισθήματα κ.λπ.
Επεξεργασία πληροφοριών στο οπτικό σύστημα
[επεξεργασία]Εξοπλισμένοι με μια πιο σταθερή κατανόηση ορισμένων από τις πιο σημαντικές έννοιες της επεξεργασίας σήματος στο οπτικό σύστημα, η κατανόηση ή η αντίληψη των επεξεργασμένων αισθητηριακών πληροφοριών είναι το τελευταίο σημαντικό τμήμα του παζλ. Η οπτική αντίληψη είναι η διαδικασία της μετάφρασης των πληροφοριών που λαμβάνονται από τα μάτια με σκοπό την κατανόηση της εξωτερικής κατάστασης των αντικειμένων. Μας κάνει να γνωρίζουμε τον κόσμο γύρω μας και μας επιτρέπει να το καταλάβουμε καλύτερα. Με βάση την οπτική αντίληψη εξοικειωνόμαστε με πρότυπα που εφαρμόζουμε στη συνέχεια αργότερα στη ζωή και λαμβάνουμε αποφάσεις βάσει αυτών και των πληροφοριών που έχουμε λάβει. Με άλλα λόγια, η επιβίωσή μας εξαρτάται από την αντίληψη. Το πεδίο της οπτικής αντίληψης χωρίστηκε σε διαφορετικά υποπεδία, λόγω του γεγονότος ότι η επεξεργασία είναι πολύ περίπλοκη και απαιτεί διαφορετικούς εξειδικευμένους μηχανισμούς για να αντιληφθεί αυτό που γίνεται ορατό. Αυτά τα υποπεδία περιλαμβάνουν: Αντίληψη χρώματος, αντίληψη κίνησης, αντίληψη βάθους και αναγνώριση προσώπου κλπ.
Βαθιά ιεραρχίες στον πρωτεύοντα οπτικό φλοιό
[επεξεργασία]
Παρά την συνεχώς αυξανόμενη υπολογιστική ισχύ των ηλεκτρονικών συστημάτων, υπάρχουν ακόμα πολλές δραστηριότητες όπου τα ζώα και οι άνθρωποι είναι εξαιρετικά ανώτεροι από τους ηλεκτρονικούς υπολογιστές – μια από αυτές είναι η αντίληψη και η συμφιλίωση των πληροφοριών. Ο κλασικός υπολογιστής, είτε αυτός του τηλεφώνου σας είτε ένας υπερυπολογιστής που αναλαμβάνει ολόκληρο το δωμάτιο, είναι κατ 'ουσία μια αριθμομηχανή. Μπορεί να εκτελέσει έναν απίστευτο αριθμό υπολογισμών σε ένα ελάχιστο χρονικό διάστημα. Αυτό που του λείπει είναι η δημιουργία αφαιρέσεων των πληροφοριών με τις οποίες συνεργάζεται. Αν προσαρμόζετε μια φωτογραφική μηχανή στον υπολογιστή σας, η εικόνα που "αντιλαμβάνεται" είναι απλά ένα πλέγμα εικονοστοιχείων, μια δισδιάστατη σειρά αριθμών. Ένας άνθρωπος θα αναγνωρίσει αμέσως τη γεωμετρία της σκηνής, τα αντικείμενα στην εικόνα, και ίσως ακόμη και το πλαίσιο του τι συμβαίνει. Αυτή η ικανότητα μας παρέχεται από αποκλειστικά βιολογικά μηχανήματα - το οπτικό σύστημα του εγκεφάλου. Επεξεργάζεται όλα όσα βλέπουμε με ιεραρχικό τρόπο, ξεκινώντας από τα απλούστερα χαρακτηριστικά της εικόνας σε πιο πολύπλοκα μέχρι την ταξινόμηση των αντικειμένων σε κατηγορίες. Ως εκ τούτου το οπτικό σύστημα λέγεται ότι έχει βαθιά ιεραρχία. Η βαθιά ιεραρχία του οπτικού συστήματος πρωτεύων έχει εμπνεύσει τους επιστήμονες υπολογιστών να δημιουργήσουν μοντέλα τεχνητών νευρωνικών δικτύων που θα διαθέτουν επίσης πολλά επίπεδα όπου κάθε ένα από αυτά δημιουργεί υψηλότερες γενικεύσεις των δεδομένων εισόδου.
Περίπου το ήμισυ του ανθρώπινου νεοκαρτέρου είναι αφιερωμένο στην όραση. Η επεξεργασία των οπτικών πληροφοριών συμβαίνει σε τουλάχιστον 10 λειτουργικά επίπεδα. Οι νευρώνες στις πρώιμες οπτικές περιοχές εξάγουν απλά χαρακτηριστικά εικόνας σε μικρές τοπικές περιοχές του οπτικού χώρου. Καθώς οι πληροφορίες μεταδίδονται σε υψηλότερες οπτικές περιοχές, οι νευρώνες ανταποκρίνονται σε όλο και πιο περίπλοκα χαρακτηριστικά. Με υψηλότερα επίπεδα επεξεργασίας πληροφοριών οι παραστάσεις γίνονται πιο αμετάβλητες - λιγότερο ευαίσθητες στο ακριβές μέγεθος, περιστροφή ή θέση του χαρακτηριστικού. Επιπλέον, αυξάνεται το μέγεθος του δεκτικού πεδίου των νευρώνων σε υψηλότερες οπτικές περιοχές, γεγονός που υποδηλώνει ότι προσαρμόζονται σε περισσότερο παγκόσμια χαρακτηριστικά εικόνας. Αυτή η ιεραρχική δομή επιτρέπει την αποτελεσματική υπολογιστική - διαφορετικές υψηλότερες οπτικές περιοχές μπορούν να χρησιμοποιούν τις ίδιες πληροφορίες που υπολογίζονται στις χαμηλότερες περιοχές. Η γενική περιγραφή της σκηνής που γίνεται στις πρώιμες οπτικές περιοχές χρησιμοποιείται από άλλα μέρη του εγκεφάλου για να ολοκληρώσει διάφορες διαφορετικές εργασίες, όπως αναγνώριση και κατηγοριοποίηση αντικειμένων, πιάσιμο, χειρισμό, σχεδιασμός κτλ.
Υποφλοιώδης όραση
[επεξεργασία]Η νευρωνική επεξεργασία των οπτικών πληροφοριών αρχίζει ήδη πριν από οποιαδήποτε από τις δομές του φλοιού. Οι φωτοϋποδοχείς στον αμφιβληστροειδή ανιχνεύουν το φως και στέλνουν σήματα στα κύτταρα γαγγλίων του αμφιβληστροειδούς. Το μέγεθος του δεκτικού χώρου ενός φωτοϋποδοχέα είναι ένα 100ο του ενός βαθμού (ένα μεγάλο, δεκτικό πεδίο είναι περίπου το μέγεθος του αντίχειρά σας, όταν το χέρι σας τεντώνεται μπροστά σας). Ο αριθμός των εισροών σε ένα κύτταρο γαγγλίων και συνεπώς το μέγεθος του δεκτικού του πεδίου εξαρτάται από τη θέση - στο κέντρο του αμφιβληστροειδούς λαμβάνει σήματα από μόλις πέντε υποδοχείς, ενώ στην περιφέρεια ένα μόνο κύτταρο μπορεί να έχει αρκετές χιλιάδες εισόδους. Αυτό σημαίνει ότι η υψηλότερη χωρική ανάλυση βρίσκεται στο κέντρο του αμφιβληστροειδούς, που ονομάζεται επίσης και βοθρίο. Λόγω αυτού του πρωτεύοντος ιδιοκτησίας κατέχει ένα μηχανισμό ελέγχου βλέμματος που κατευθύνει την όραση, έτσι ώστε τα χαρακτηριστικά ενδιαφέροντος να προβάλλονται πάνω στο βοθρίο.
Τα γαγγλιακά κύτταρα είναι επιλεκτικά συντονισμένα στην ανίχνευση διαφόρων χαρακτηριστικών της εικόνας, όπως η αντίθεση φωτεινότητας, η χρωματική αντίθεση και η κατεύθυνση και η ταχύτητα κίνησης. Όλες αυτές οι λειτουργίες είναι οι κύριες πληροφορίες που χρησιμοποιούνται περαιτέρω στον αγωγό επεξεργασίας. Αν υπάρχουν οπτικά ερεθίσματα που δεν ανιχνεύονται από τα κύτταρα των γαγγλίων, τότε δεν είναι διαθέσιμα για οποιαδήποτε φλοιώδη οπτική περιοχή.
Τα κύτταρα των γαγγλίων προβάλλουν σε μια περιοχή στον θάλαμο που ονομάζεται πλευρικός πυρήνας γονιδιώματος (LGN), ο οποίος με τη σειρά του μεταδίδει τα σήματα στον φλοιό. Δεν υπάρχει αξιόλογος υπολογισμός που να είναι γνωστός στο LGN - υπάρχει σχεδόν μια αντιστοιχία ενός προς ένα μεταξύ του γαγγλίου του αμφιβληστροειδούς και των κυττάρων LGN. Ωστόσο, μόνο το 5% των εισροών στην LGN προέρχεται από τον αμφιβληστροειδή - όλες οι άλλες εισροές είναι προβολές φλοιώδους ανάδρασης. Αν και το οπτικό σύστημα θεωρείται συχνά σύστημα προώθησης, οι επαναλαμβανόμενες συνδέσεις ανάδρασης καθώς και οι πλευρικές συνδέσεις είναι ένα κοινό χαρακτηριστικό που παρατηρείται σε όλο τον οπτικό φλοιό. Ο ρόλος της ανατροφοδότησης δεν είναι ακόμη πλήρως κατανοητός, αλλά προτείνεται να αποδοθεί σε διαδικασίες όπως η προσοχή, η προσδοκία, η φαντασία και η συμπλήρωση των πληροφοριών που λείπουν.
Οφθαλμική όραση
[επεξεργασία]
Ο οπτικός φλοιός μπορεί να χωριστεί σε τρία μεγάλα τμήματα - το ινιακό τμήμα που λαμβάνει εισροή από το LGN και έπειτα στέλνει σήματα στα ραχιαία και κοιλιακά ρεύματα. Το περιθωριακό μέρος περιλαμβάνει τις περιοχές V1-V4 και MT, οι οποίες επεξεργάζονται διάφορες πτυχές της οπτικής πληροφορίας και δημιουργούν μια γενική απεικόνιση της σκηνής. Η ραχιαία δίοδος συμμετέχει στην ανάλυση του χώρου και του σχεδιασμού δράσης. Η κοιλιακή δίοδος εμπλέκεται στην αναγνώριση και κατηγοριοποίηση αντικειμένων.

Το V1 είναι η πρώτη φλοιώδης περιοχή που επεξεργάζεται οπτικές πληροφορίες. Είναι ευαίσθητο στις άκρες, παράλληλες γραμμές, το τέλος των γραμμών, τις κινήσεις, το χρώμα και τις ανισότητες (γωνιακή διαφορά μεταξύ των προβολών ενός σημείου στον αριστερό και στον δεξιό αμφιβληστροειδή). Το πιο άμεσο παράδειγμα της ιεραρχικής επεξεργασίας από τη βάση προς τα πάνω είναι ο γραμμικός συνδυασμός των εισροών από διάφορα κύτταρα των γαγγλιοκυττάρων με κεντρικά δεκτικά πεδία για να δημιουργηθεί μια αναπαράσταση ενός ραβδίου. Αυτό γίνεται από τα απλά κύτταρα του V1 και περιγράφηκε για πρώτη φορά από τους εξέχοντες νευροεπιστήμονες Hubel και Wiesel. Αυτός ο τύπος ενοποίησης πληροφοριών υποδηλώνει ότι τα απλά κύτταρα είναι ευαίσθητα στην ακριβή θέση της ραβδίου και έχουν ένα σχετικά μικρό δεκτικό πεδίο. Τα σύνθετα κύτταρα του V1 λαμβάνουν εισροές από τα απλά κύτταρα και ενώ ανταποκρίνονται επίσης σε γραμμικά προσανατολισμένα μοτίβα, δεν είναι ευαίσθητα στην ακριβή θέση της ραβδίου και έχουν μεγαλύτερο πεδίο δεκτικότητας. Ο υπολογισμός που υπάρχει σε αυτό το βήμα θα μπορούσε να είναι μια λειτουργία τύπου MAX που παράγει αποκρίσεις παρόμοιες σε πλάτος με τις μεγαλύτερες από τις απαντήσεις που αφορούν τα μεμονωμένα ερεθίσματα. Ορισμένα απλά και πολύπλοκα κύτταρα μπορούν επίσης να ανιχνεύσουν το άκρο μιας ραβδίου και ένα κλάσμα των κυττάρων V1 είναι επίσης ευαίσθητα στην τοπική κίνηση μέσα στα αντίστοιχα δεκτικά πεδία.

Η περιοχή V2 διαθέτει πιο εκλεπτυσμένη αναπαράσταση περιγράμματος, όπως περιγράμματα που ορίζονται από την υφή, παραπλανητικά περιγράμματα και περιγράμματα ποθ συμπεριλαμωάνουν σύνορα. Το V2 επίσης βασίζεται στην ανίχνευση απόλυτης ανισότητας σε V1 και χαρακτηρίζει κύτταρα που είναι ευαίσθητα σε σχετικές ανισότητες που είναι η διαφορά μεταξύ των απόλυτων ανισοτήτων δύο σημείων στο διάστημα. Η περιοχή V4 λαμβάνει εισροές από το V2 και την περιοχή V3, αλλά πολύ λίγα είναι γνωστά για τον υπολογισμό που λαμβάνει χώρα στο V3. Η περιοχή V4 διαθέτει νευρώνες που είναι ευαίσθητες σε περιγράμματα με διαφορετική καμπυλότητα και κορυφές με συγκεκριμένες γωνίες. Ένα άλλο σημαντικό χαρακτηριστικό είναι η κωδικοποίηση για την ανθεκτική στη φωτεινότητα απόχρωση. Αυτό έρχεται σε αντίθεση με την V1 όπου οι νευρώνες ανταποκρίνονται στην αντίθεση του χρώματος κατά μήκος των δύο βασικών αξόνων (κόκκινο-πράσινο και κίτρινο-μπλε) αντί του πραγματικού χρώματος. Το V4 εξέρχεται περαιτέρω στο κοιλιακό ρεύμα, στον φλεβοτονωτικός φλοιό (IT), ο οποίος έχει αποδειχθεί μέσω μελετών των αλλοιώσεων ότι είναι απαραίτητος για τη διάκριση αντικειμένων.
Ο φλεβοτονωτικός φλοιός: διάκριση αντικειμένων
[επεξεργασία]
Ο φλεβοτονωτικός φλοιός (Inferior temporal cortex, ΙΤ) χωρίζεται σε δύο περιοχές: TEO και TE. Το πεδίο TEO ενσωματώνει πληροφορίες σχετικά με τα σχήματα και τις σχετικές θέσεις πολλαπλών στοιχείων περιγράμματος και χαρακτηρίζει κυρίως κύτταρα τα οποία ανταποκρίνονται σε απλούς συνδυασμούς χαρακτηριστικών. Το μέγεθος του δεκτικού πεδίου των νευρώνων ΤΕΟ είναι περίπου 3-5 μοίρες. Η περιοχή ΤΕ διαθέτει κύτταρα με σημαντικά μεγαλύτερα δεκτικά πεδία (10-20 μοίρες) τα οποία ανταποκρίνονται σε πρόσωπα, χέρια και σύνθετες διαμορφώσεις χαρακτηριστικών. Τα κύτταρα στην ΤΕ ανταποκρίνονται σε οπτικά χαρακτηριστικά που είναι μια απλούστερη γενίκευση του αντικειμένου ενδιαφέροντος, αλλά πιο περίπλοκα από απλές ραβδίους ή κηλίδες. Αυτό απεδείχθη χρησιμοποιώντας μία μέθοδο μείωσης ερεθίσματος από τους Tanaka κ.ά. όπου πρώτα μετράται μια απόκριση σε ένα αντικείμενο και στη συνέχεια το αντικείμενο αντικαθίσταται από απλούστερες αναπαραστάσεις έως ότου το κρίσιμο χαρακτηριστικό στο οποίο αντιδρούν οι νευρώνες ΤΕ περιορίζεται.
Φαίνεται ότι οι νευρώνες στον ΙΤ συναρμολογούν διάφορα χαρακτηριστικά μεσαίας πολυπλοκότητας από χαμηλότερα επίπεδα στο κοιλιακό ρεύμα για να διαμορφώσουν μοντέλα αντικειμένων. Οι νευρώνες στην ΤΕ που είναι επιλεκτικοί σε συγκεκριμένα αντικείμενα πρέπει να πληρούν δύο φαινομενικά αντιφατικές απαιτήσεις - επιλεκτικότητα και αμεταβλητότητα. Πρέπει να κάνουν διάκριση μεταξύ διαφορετικών αντικειμένων με τη βοήθεια της ευαισθησίας στα χαρακτηριστικά των εικόνων του αμφιβληστροειδούς. Ωστόσο, το ίδιο αντικείμενο μπορεί να αντιμετωπιστεί από διαφορετικές γωνίες και αποστάσεις σε διαφορετικές συνθήκες φωτισμού, αποδίδοντας εξαιρετικά ανόμοιες εικόνες αμφιβληστροειδούς του ίδιου αντικειμένου. Για να αντιμετωπιστούν όλες αυτές οι εικόνες ως ισοδύναμες, πρέπει να παραχθούν αμετάβλητα χαρακτηριστικά τα οποία είναι ανθεκτικά σε ορισμένους μετασχηματισμούς, όπως αλλαγές στη θέση, φωτισμό, μέγεθος στον αμφιβληστροειδή κλπ. Οι νευρώνες στην περιοχή ΤΕ δείχνουν αμετάβλητη θέση και μέγεθος καθώς και μερική απόφραξη , θέση στον χώρο και κατεύθυνση φωτισμού. Η περιστροφή σε βάθος χώρου έχει αποδειχθεί ότι έχει την ασθενέστερη μεταβλητότητα, με εξαίρεση εαν το αντικείμενο είναι ανθρώπινο πρόσωπο.
Οι κατηγορίες αντικειμένων δεν υπάρχουν ακόμη απόλυτα στην περιοχή ΤΕ - ένας νευρώνας μπορεί τυπικά να ανταποκρίνεται σε πολλά αλλά όχι όλα τα υποδείγματα της ίδιας κατηγορίας (π.χ. εικόνες δέντρων) και μπορεί επίσης να ανταποκρίνεται σε υποδείγματα διαφορετικών κατηγοριών (π.χ. δέντρα και μη- δέντρα). Η αναγνώριση αντικειμένων και η ταξινόμηση πιθανότατα περιλαμβάνει δειγματοληψία από μεγαλύτερο πληθυσμό νευρώνων ΤΕ καθώς και λήψη εισερχομένων από πρόσθετες περιοχές του εγκεφάλου, π.χ. εκείνες που είναι υπεύθυνες για την κατανόηση του πλαισίου της σκηνής. Πρόσφατα πειράματα ανάγνωσης απέδειξαν ότι οι στατιστικοί ταξινομητές (π.χ. μηχανές φορέων υποστήριξης) μπορούν να εκπαιδευτούν για την ταξινόμηση αντικειμένων με βάση τις αποκρίσεις ενός μικρού αριθμού νευρώνων ΤΕ. Ως εκ τούτου, ένας πληθυσμός των νευρώνων ΤΕ μπορεί κατ 'αρχήν να επισημάνει αξιόπιστα κατηγορίες αντικειμένων με τη συνδυασμένη δραστηριότητα τους. Είναι ενδιαφέρον ότι υπάρχουν επίσης αναφορές για εξαιρετικά επιλεκτικούς νευρώνες στον μεσαίο κροταφικό λοβό που ανταποκρίνονται σε πολύ συγκεκριμένες ενδείξεις, π.χ. στον πύργο της Πίζας σε διαφορετικές εικόνες ή στο πρόσωπο ενός συγκεκριμένου προσώπου.
Μάθηση στο οπτικό σύστημα
[επεξεργασία]Η εκμάθηση μπορεί να μεταβάλει την εκλεκτικότητα οπτικών χαρακτηριστικών των νευρώνων, με αποτέλεσμα να μαθαίνει να γίνεται ισχυρότερη σε υψηλότερα ιεραρχικά επίπεδα. Δεν υπάρχουν γνωστά στοιχεία για τη μάθηση στον αμφιβληστροειδή και επίσης οι χάρτες προσανατολισμού στο V1 φαίνεται να είναι γενικά γενετικά προκαθορισμένοι. Ωστόσο, η πρακτική αναγνώριση προσανατολισμού βελτιώνει την κωδικοποίηση προσανατολισμού στους νευρώνες V1, αυξάνοντας την κλίση της καμπύλης συντονισμού. Παρόμοια αλλά μεγαλύτερα αποτελέσματα έχουν παρατηρηθεί στο V4. Στην περιοχή της ΤΕ σχετικά μικρή οπτική κατάρτιση έχει αξιοσημείωτες φυσιολογικές επιδράσεις στην οπτική αντίληψη, σε ένα μόνο επίπεδο κυττάρων καθώς και στο fMRI. Για παράδειγμα, η μορφοποίηση δύο αντικειμένων μεταξύ τους αυξάνει την αντιληπτή ομοιότητά τους. Συνολικά, φαίνεται ότι ο ομοιόμορφος οπτικός φλοιός των ενηλίκων είναι αισθητά πλαστικός και το επίπεδο πλαστικότητας μπορεί να αυξηθεί σημαντικά, π.χ. χορηγώντας συγκεκριμένα φάρμακα ή ζώντας σε ένα εμπλουτισμένο περιβάλλον.
Bαθιά Νευρωνικά Δίκτυα
[επεξεργασία]Οπως συμβαίνει με τη βαθιά ιεραρχία του πρωτεύοντος οπτικού συστήματος, οι αρχιτεκτονικές βαθιάς εκμάθησης επιχειρούν να μοντελοποιήσουν υψηλού επιπέδου απλοποιήσεις της εισαγωγής δεδομένων χρησιμοποιώντας πολλαπλά επίπεδα μη γραμμικών μετασχηματισμών. Το μοντέλο που πρότειναν οι Hubel και Wiesel, όπου οι πληροφορίες ενσωματώνονται και πολλαπλασιάζονται σε μια αλληλουχία από τον αμφιβληστροειδή και το LGN σε απλά κύτταρα και σύνθετα κύτταρα στην V1, ενέπνευσαν τη δημιουργία μιας από τις πρώτες αρχιτεκτονικές βαθιάς εκμάθησης, το neocognitron - ένα μοντέλο πολλαπλών στρωμάτων τεχνητού νευρικού δικτύου. Χρησιμοποιήθηκε για διαφορετικές εργασίες αναγνώρισης προτύπων, συμπεριλαμβανομένης της αναγνώρισης χειρόγραφων χαρακτήρων. Ωστόσο, χρειάστηκε πολύς χρόνος για την κατάρτιση του δικτύου (σε επίπεδο ημερών) και από την έναρξή του στη δεκαετία του 1980 η βαθιά εκμάθηση δεν έλαβε μεγάλη προσοχή μέχρι τα μέσα της δεκαετίας του 2000 με την αφθονία των ψηφιακών δεδομένων και την εφεύρεση αλγορίθμων ταχείας εκμάθησης. Τα βαθιά νευρωνικά δίκτυα αποδείχθηκαν πολύ αποτελεσματικά σε καθήκοντα που οχι και τόσο πολύ καιρό πριν ήταν δυνατά μόνο για τον άνθρωπο, όπως η αναγνώριση των προσώπων συγκεκριμένων ανθρώπων στις φωτογραφίες, η κατανόηση του ανθρώπινου λόγου (σε κάποιο βαθμό) και η μετάφραση κειμένου από ξένες γλώσσες . Επιπλέον, έχουν αποβεί σε μεγάλη βοήθεια στη βιομηχανία και την επιστήμη για την αναζήτηση πιθανών υποψήφιων φαρμάκων, χαρτογράφηση πραγματικών νευρωνικών δικτύων στον εγκέφαλο και πρόβλεψη των λειτουργιών των πρωτεϊνών. Πρέπει να σημειωθεί ότι η βαθιά εκμάθηση είναι μόνο πολύ χαλαρά εμπνευσμένη από τον εγκέφαλο και είναι πολύ περισσότερο ένα επίτευγμα του πεδίου της επιστήμης των υπολογιστών / μηχανικής μάθησης παρά της νευροεπιστήμης. Οι βασικές ομοιότητες είναι ότι τα βαθιά νευρωνικά δίκτυα αποτελούνται από μονάδες που ενσωματώνουν εισόδους πληροφοριών με μη γραμμικό τρόπο (νευρώνες) και στέλνουν σήματα μεταξύ τους (συνάψεις) και ότι υπάρχουν διαφορετικά επίπεδα όλο και πιο αφηρημένων αναπαραστάσεων των δεδομένων. Οι αλγόριθμοι μάθησης και οι μαθηματικές περιγραφές των «νευρώνων» που χρησιμοποιούνται στη βαθιά εκμάθηση είναι πολύ διαφορετικοί από τις πραγματικές διαδικασίες που λαμβάνουν χώρα στον εγκέφαλο. Ως εκ τούτου, η έρευνα στη βαθιά εκμάθηση, δίνοντας μια τεράστια ώθηση σε μια πιο εξελιγμένη τεχνητή νοημοσύνη, μπορεί να δώσει περιορισμένες γνώσεις για τον εγκέφαλο.
Bιβλιογραφικές Aναφορές
[επεξεργασία]- Έγγραφα σχετικά με τις βαθιές ιεραρχίες στο οπτικό σύστημα
- (August 2013) "Deep Hierarchies in the Primate Visual Cortex: What Can We Learn for Computer Vision?". IEEE Transactions on Pattern Analysis and Machine Intelligence 35 (8): 1847–1871. DOI:10.1109/TPAMI.2012.272.
- (1 November 1999) "{{{title}}}". Nature Neuroscience 2 (11): 1019–1025. DOI:doi:10.1038/14819.
- Πείραμα μείωσης τόνωσης
- (March 1996) "Inferotemporal Cortex and Object Vision". Annual Review of Neuroscience 19 (1): 109–139. DOI:10.1146/annurev.ne.19.030196.000545.
- Στοιχεία μάθησης στο οπτικό σύστημα
- (23 September 2010) "Unsupervised Natural Visual Experience Rapidly Reshapes Size-Invariant Object Representation in Inferior Temporal Cortex". Neuron 67 (6): 1062–1075. DOI:10.1016/j.neuron.2010.08.029.
- (14 June 2006) "Learning to See the Difference Specifically Alters the Most Informative V4 Neurons". Journal of Neuroscience 26 (24): 6589–6602. DOI:10.1523/JNEUROSCI.0457-06.2006.
- (2 August 2001) "Practising orientation identification improves orientation coding in V1 neurons.". Nature 412 (6846): 549-53.
- Μια πρόσφατη και προσβάσιμη επισκόπηση του status quo της έρευνας βαθιάς μάθησης
- (8 January 2014) "Computer science: The learning machines". Nature 505 (7482): 146–148. DOI:10.1038/505146a.
Αντίληψη κίνησης
[επεξεργασία]Η αντίληψη κίνησης είναι η διαδικασία υπολογισμού της ταχύτητας και της κατεύθυνσης των κινούμενων αντικειμένων. Η περιοχή V5 στον άνθρωπο και στην περιοχή MT (Middle Temporal) στα πρωτεύοντα θηλαστικά είναι υπεύθυνη για την φλοιική αντίληψη της κίνησης. Η περιοχή V5 είναι μέρος του εξωστρεφούς φλοιού, που είναι η περιοχή στην ινιακή περιοχή του εγκεφάλου δίπλα στον πρωτογενή οπτικό φλοιό. Η λειτουργία της Περιοχής V5 είναι να ανιχνεύσει την ταχύτητα και την κατεύθυνση των οπτικών ερεθισμάτων και να ενσωματώσει τα τοπικά οπτικά σήματα κίνησης σε παγκόσμια κίνηση. Η περιοχή V1 ή πρωτογενής οπτικός φλοιός βρίσκεται στον ινιακό λοβό του εγκεφάλου και στα δύο ημισφαίρια. Διαχειρίζεται το πρώτο στάδιο της φλοιώδους επεξεργασίας των οπτικών πληροφοριών. Αυτή η περιοχή περιέχει έναν πλήρη χάρτη του οπτικού πεδίου που καλύπτεται από τα μάτια. Η διαφορά μεταξύ της περιοχής V5 και της περιοχής V1 (Primary Visual Cortex) είναι ότι η περιοχή V5 μπορεί να ενσωματώσει την κίνηση τοπικών σημάτων ή μεμονωμένων τμημάτων ενός αντικειμένου σε μια συνολική κίνηση ενός ολόκληρου αντικειμένου. Η περιοχή V1, από την άλλη πλευρά, αποκρίνεται στην τοπική κίνηση που συμβαίνει εντός του δεκτικού πεδίου. Οι εκτιμήσεις από αυτούς τους πολλούς νευρώνες είναι ενσωματωμένες στην περιοχή V5.
Η κίνηση ορίζεται ως αλλαγές στον φωτισμό του αμφιβληστροειδούς στο διάστημα και στο χρόνο. Τα σήματα κίνησης ταξινομούνται σε κινήσεις πρώτης τάξης και κινήσεις δεύτερης τάξης. Αυτοί οι τύποι κίνησης περιγράφονται εν συντομία στις επόμενες παραγράφους.

Η αντίληψη κίνησης πρώτης τάξης αναφέρεται στην κίνηση που γίνεται αντιληπτή όταν δύο ή περισσότερα οπτικά ερεθίσματα ενεργοποιούν και απενεργοποιούν την πάροδο του χρόνου και παράγουν διαφορετικές αντιλήψεις κίνησης. Η κίνηση της πρώτης τάξης ονομάζεται επίσης «φαινομενική κίνηση» και χρησιμοποιείται στην τηλεόραση και στην ταινία. Ένα παράδειγμα αυτού είναι το «κίνημα της βήτα», το οποίο είναι μια ψευδαίσθηση στην οποία οι σταθερές εικόνες φαίνεται να κινούνται, παρόλο που δεν κινούνται Η οπτική αυτή ψευδαίσθηση συμβαίνει επειδή το ανθρώπινο οπτικό νεύρο αποκρίνεται σε αλλαγές φωτός σε δέκα κύκλους ανά δευτερόλεπτο, έτσι ώστε οποιαδήποτε αλλαγή να είναι ταχύτερη από αυτή την ταχύτητα θα εγγραφεί ως κίνηση συνεχούς λειτουργίας και όχι ως ξεχωριστές εικόνες.
Η κίνηση δεύτερης τάξης αναφέρεται στην κίνηση που εμφανίζεται όταν ένα μετακινούμενο περίγραμμα ορίζεται από την αντίθεση, την υφή, την τρεμόπαιξη ή κάποια άλλη ποιότητα που δεν έχει ως αποτέλεσμα την αύξηση της φωτεινότητας ή της κίνησης της εικόνας. Τα στοιχεία δείχνουν ότι η έγκαιρη επεξεργασία της κίνησης πρώτης τάξης και της κίνησης δεύτερης τάξης πραγματοποιείται με ξεχωριστές διαδρομές. Οι μηχανισμοί δεύτερης τάξης έχουν φτωχότερη χρονική ανάλυση και είναι χαμηλής διέλευσης από την άποψη της κλίμακας χωρικών συχνοτήτων στις οποίες ανταποκρίνονται. Η κίνηση δεύτερης τάξης παράγει μια πιο αδύναμη κίνηση. Τα σήματα πρώτης και δεύτερης τάξης συνδυάζονται στο V5.
Σε αυτό το κεφάλαιο θα αναλύσουμε τις έννοιες Ανίχνευση κίνησης και ανάλυσης κινήσεων και θα εξηγήσουμε τον λόγο για τον οποίο οι όροι αυτοί δεν θα πρέπει να χρησιμοποιούνται εναλλακτικά. Θα αναλύσουμε τους μηχανισμούς με τους οποίους γίνεται αντιληπτή η κίνηση όπως οι αισθητήρες κίνησης και η παρακολούθηση χαρακτηριστικών. Υπάρχουν τρία κύρια θεωρητικά μοντέλα που προσπαθούν να περιγράψουν τη λειτουργία των νευρωνικών αισθητήρων κίνησης. Έχουν διεξαχθεί πειραματικές δοκιμές για να επιβεβαιωθεί η ακρίβεια αυτών των μοντέλων. Δυστυχώς, τα αποτελέσματα αυτών των δοκιμών είναι ασαφή και μπορεί να ειπωθεί ότι κανένα από αυτά τα μοντέλα δεν περιγράφει πλήρως τη λειτουργία των αισθητήρων κίνησης. Ωστόσο, κάθε ένα από αυτά τα μοντέλα προσομοιώνει ορισμένα χαρακτηριστικά των αισθητήρων κίνησης. Περιγράφονται μερικές ιδιότητες αυτών των αισθητήρων. Τέλος, αυτό το κεφάλαιο παρουσιάζει κάποιες ψευδαισθήσεις κίνησης, που δείχνουν ότι η αίσθηση κίνησης μπορεί να παραπλανηθεί από στατικούς εξωτερικούς παράγοντες που διεγείρουν τους αισθητήρες κίνησης με τον ίδιο τρόπο όπως και η κίνηση.
Ανάλυση κίνησης και αντίληψη κίνησης
[επεξεργασία]Οι έννοιες της ανάλυσης κίνησης και της αντίληψης κίνησης συχνά συγχέονται ως εναλλάξιμες. Η αντίληψη κίνησης και η ανάλυση κίνησης είναι σημαντικές μεταξύ τους, αλλά δεν είναι οι ίδιες. Ανάλυση κίνησης αναφέρεται στους μηχανισμούς στους οποίους επεξεργάζονται τα σήματα κίνησης. Με παρόμοιο τρόπο, όπου η αντίληψη κίνησης δεν εξαρτάται απαραίτητα από τα σήματα που δημιουργούνται από την κίνηση εικόνων στον αμφιβληστροειδή, η ανάλυση κίνησης μπορεί να οδηγήσει ή να μην οδηγήσει σε αντίληψη κίνησης. Ένα παράδειγμα αυτού του φαινομένου είναι το Vection, το οποίο συμβαίνει όταν ένα άτομο αντιλαμβάνεται ότι κινείται όταν είναι ακίνητο, αλλά το αντικείμενο που παρατηρεί κινείται. Η εντολή δείχνει ότι η κίνηση ενός αντικειμένου μπορεί να αναλυθεί, αν και δεν θεωρείται κίνηση που προέρχεται από το αντικείμενο. Αυτός ο ορισμός της ανάλυσης κίνησης υποδηλώνει ότι η κίνηση είναι μια βασική ιδιότητα εικόνας. Στο οπτικό πεδίο, αναλύεται σε κάθε σημείο. Τα αποτελέσματα αυτής της ανάλυσης χρησιμοποιούνται για την εξαγωγή αντιληπτικών πληροφοριών.
Η αντίληψη κίνησης αναφέρεται στη διαδικασία απόκτησης αντιληπτικής γνώσης σχετικά με την κίνηση αντικειμένων και επιφανειών σε μια εικόνα. Η κίνηση γίνεται αντιληπτή είτε από ευαίσθητους τοπικούς αισθητήρες στον αμφιβληστροειδή είτε από παρακολούθηση χαρακτηριστικών. Οι αισθητήρες τοπικής κίνησης είναι εξειδικευμένοι νευρώνες ευαίσθητοι στην κίνηση και είναι ανάλογοι με εξειδικευμένους αισθητήρες για το χρώμα. Η παρακολούθηση χαρακτηριστικών είναι ένας έμμεσος τρόπος για να αντιληφθεί κανείς την κίνηση και συνίσταται στην υποκίνηση κίνησης από αλλαγές στην αμφιβληστροειδική θέση αντικειμένων με την πάροδο του χρόνου. Αναφέρεται επίσης ως ανάλυση κίνησης τρίτης τάξης. Η παρακολούθηση χαρακτηριστικών λειτουργεί εστιάζοντας την προσοχή σε ένα συγκεκριμένο αντικείμενο και παρατηρώντας τον τρόπο με τον οποίο η θέση του έχει αλλάξει με την πάροδο του χρόνου.
Αισθητήρες κίνησης
[επεξεργασία]Η ανίχνευση της κίνησης είναι το πρώτο στάδιο της οπτικής επεξεργασίας και συμβαίνει χάρη στις εξειδικευμένες νευρικές διεργασίες, οι οποίες ανταποκρίνονται στις πληροφορίες σχετικά με τις τοπικές αλλαγές της έντασης των εικόνων με την πάροδο του χρόνου. Η κίνηση ανιχνεύεται ανεξάρτητα από άλλες ιδιότητες εικόνας σε όλες τις θέσεις της εικόνας. Έχει αποδειχθεί ότι υπάρχουν αισθητήρες κίνησης και λειτουργούν τοπικά σε όλα τα σημεία της εικόνας. Οι αισθητήρες κίνησης είναι εξειδικευμένοι νευρωνικοί αισθητήρες τοποθετημένοι στον αμφιβληστροειδή που είναι ικανοί να ανιχνεύουν μια κίνηση που παράγεται από δύο μικρές και μικρές φωτεινές αναλαμπές που είναι τόσο κοντά μεταξύ τους ώστε να μην μπορούν να ανιχνευθούν από την παρακολούθηση χαρακτηριστικών. Υπάρχουν τρία βασικά μοντέλα που προσπαθούν να περιγράψουν τον τρόπο με τον οποίο λειτουργούν αυτοί οι εξειδικευμένοι αισθητήρες. Αυτά τα μοντέλα είναι ανεξάρτητα το ένα από το άλλο και προσπαθούν να διαμορφώσουν συγκεκριμένα χαρακτηριστικά της αντίληψης κίνησης. Παρόλο που δεν υπάρχουν επαρκή στοιχεία για να υποστηριχθεί ότι κάποιο από αυτά τα μοντέλα αντιπροσωπεύει τον τρόπο που το οπτικό σύστημα (αισθητήρες κίνησης ιδιαίτερα) αντιλαμβάνεται την κίνηση, εξακολουθούν να διαμορφώνουν σωστά ορισμένες λειτουργίες αυτών των αισθητήρων.
Ο ανιχνευτής Reichardt
[επεξεργασία]
Ο ανιχνευτής Reichardt χρησιμοποιείται για να υποδείξει πώς οι αισθητήρες
κίνησης ανταποκρίνονται στα σήματα κίνησης πρώτης τάξης. Όταν ένα
αντικείμενο μετακινείται από το σημείο Α στο οπτικό πεδίο στο σημείο Β,
παράγονται δύο σήματα: ένα πριν ξεκινήσει η κίνηση και ένα άλλο μετά την
ολοκλήρωση της κίνησης. Αυτό το μοντέλο αντιλαμβάνεται αυτή την κίνηση
ανιχνεύοντας αλλαγές φωτεινότητας σε ένα σημείο στον αμφιβληστροειδή και
συσχετίζοντάς το με μια αλλαγή φωτεινότητας σε άλλο σημείο κοντά μετά από
μια σύντομη καθυστέρηση. Ο ανιχνευτής Reichardt λειτουργεί με βάση την αρχή
της συσχέτισης (στατιστική σχέση που περιλαμβάνει εξάρτηση). Ερμηνεύει ένα
σήμα κίνησης με χωροχρονική συσχέτιση σημάτων φωτεινότητας στα γειτονικά
σημεία. Χρησιμοποιεί το γεγονός ότι δύο δεκτικά πεδία σε διαφορετικά σημεία
στην τροχιά ενός κινούμενου αντικειμένου λαμβάνουν μια χρονικά
μετατοπισμένη έκδοση του ίδιου σήματος - ένα μοτίβο φωτεινότητας κινείται
κατά μήκος ενός άξονα και το σήμα σε ένα σημείο του άξονα είναι μια χρονικά
μετατοπισμένη έκδοση του ένα προηγούμενο σήμα στον άξονα. Το μοντέλο
ανιχνευτή Reichardt διαθέτει δύο χωριστά χωριστούς ανιχνευτές γειτνίασης.
Τα σήματα εξόδου των ανιχνευτών πολλαπλασιάζονται (συσχετίζονται) με τον
ακόλουθο τρόπο: ένα σήμα πολλαπλασιασμένο με ένα δεύτερο σήμα που είναι η
χρονικά μετατοπισμένη εκδοχή του πρωτοτύπου.
Η ίδια διαδικασία
επαναλαμβάνεται αλλά στην αντίστροφη κατεύθυνση της κίνησης (το σήμα που
μετατοπίστηκε στο χρόνο γίνεται το πρώτο σήμα και το αντίστροφο). Στη
συνέχεια, λαμβάνεται η διαφορά μεταξύ αυτών των δύο πολλαπλασιασμών και το
αποτέλεσμα δίνει την ταχύτητα κίνησης. Η απόκριση του ανιχνευτή εξαρτάται
από τη φάση, την αντίθεση και την ταχύτητα του ερεθίσματος. Πολλοί
ανιχνευτές που συντονίζονται με διαφορετικές ταχύτητες είναι απαραίτητοι
για την κωδικοποίηση της πραγματικής ταχύτητας του σχεδίου. Τα πιο
συναρπαστικά πειραματικά στοιχεία για αυτό το είδος ανιχνευτή προέρχονται
από μελέτες της διάκρισης κατεύθυνσης των ελάχιστα ορατών στόχων.
Φιλτράρισμα κίνησης-ενέργειας
[επεξεργασία]Το φίλτρο ενέργειας κίνησης είναι ένα μοντέλο αισθητήρων κίνησης που βασίζεται στην αρχή των φίλτρων αμετάβλητης φάσης. Αυτό το μοντέλο δημιουργεί χωροχρονικά φίλτρα προσανατολισμένα σε χωροχρόνο ώστε να ταιριάζουν με τη δομή των κινούμενων μοτίβων. Αποτελείται από διαχωρίσιμα φίλτρα, για τα οποία τα χωρικά προφίλ παραμένουν το ίδιο σχήμα με την πάροδο του χρόνου αλλά κλιμακώνονται με την τιμή των χρονικών φίλτρων. Τα φίλτρα ενέργειας κίνησης ταιριάζουν με τη δομή των κινούμενων μοτίβων προσθέτοντας μαζί διαχωρίσιμα φίλτρα Για κάθε κατεύθυνση κίνησης δημιουργούνται δύο φίλτρα χωροχρόνου: ένα, το οποίο είναι συμμετρικό (σαν ραβδίο) και το ένα είναι ασύμμετρο (άκρο). Το άθροισμα των τετραγώνων αυτών των φίλτρων καλείται ενέργεια κίνησης. Η διαφορά στο σήμα για τις δύο κατευθύνσεις ονομάζεται ενέργεια του αντιπάλου. Αυτό το αποτέλεσμα διαιρείται στη συνέχεια από την τετραγωνική έξοδο ενός άλλου φίλτρου, το οποίο συντονίζεται σε στατική αντίθεση. Αυτή η διαίρεση γίνεται για να ληφθεί υπόψη η επίδραση της αντίθεσης στην κίνηση. Τα φίλτρα ενέργειας κίνησης μπορούν να μοντελοποιήσουν ένα αριθμό φαινομένων κίνησης, αλλά παράγουν μια ανεξάρτητη από τη φάση μέτρηση, η οποία αυξάνεται με την ταχύτητα αλλά δεν δίνει αξιόπιστη τιμή ταχύτητας.
Χωροχρονική Διαβαθμίσεις
[επεξεργασία]
Αυτό το μοντέλο αισθητήρων κίνησης αναπτύχθηκε αρχικά στον τομέα της όρασης
του υπολογιστή και βασίζεται στην αρχή ότι η αναλογία του χρονικού
παραγώγου της φωτεινότητας της εικόνας με το χωρικό παράγωγο της
φωτεινότητας της εικόνας δίνει την ταχύτητα κίνησης. Είναι σημαντικό να
σημειωθεί ότι στις κορυφές και τις καμπύλες της εικόνας, αυτό το μοντέλο
δεν θα υπολογίσει μια επαρκή απάντηση, επειδή το παράγωγο στον παρονομαστή
θα είναι μηδέν. Για την επίλυση αυτού του προβλήματος, μπορούν επίσης να
αναλυθούν τα χωρικά παράγωγα πρώτης τάξης και υψηλότερης τάξης σε σχέση με
το χώρο και το χρόνο. Οι Spatiotemporal Gradients είναι ένα καλό μοντέλο
για τον προσδιορισμό της ταχύτητας κίνησης σε όλα τα σημεία της εικόνας.
Οι αισθητήρες κίνησης είναι Προσανατολισμός-Επιλεκτικός
[επεξεργασία]Μία από τις ιδιότητες των αισθητήρων κίνησης είναι η επιλεκτικότητα προσανατολισμού, η οποία περιορίζει την ανάλυση κίνησης σε μία μόνο διάσταση. Οι αισθητήρες κίνησης μπορούν να καταγράφουν κίνηση μόνο σε μία διάσταση κατά μήκος ενός άξονα ορθογώνιου προς τον προτιμώμενο προσανατολισμό του αισθητήρα. Ένα ερέθισμα που περιέχει χαρακτηριστικά ενός μόνο προσανατολισμού μπορεί μόνο να θεωρηθεί ότι κινείται προς μια κατεύθυνση ορθή προς τον προσανατολισμό του ερεθίσματος. Τα μονοδιάστατα σήματα κίνησης δίνουν διφορούμενες πληροφορίες σχετικά με την κίνηση των δισδιάστατων αντικειμένων. Ένα δεύτερο στάδιο της ανάλυσης κίνησης είναι απαραίτητο για την επίλυση της πραγματικής κατεύθυνσης κίνησης ενός αντικειμένου ή μοτίβου 2-D. Τα σήματα κίνησης 1-D από αισθητήρες συντονισμένους σε διαφορετικούς προσανατολισμούς συνδυάζονται για να παράγουν ένα σαφές σήμα κίνησης 2-D. Η ανάλυση της κίνησης 2-D εξαρτάται από σήματα από τοπικούς ευρέως προσανατολισμένους αισθητήρες καθώς και από σήματα από στενά προσανατολισμένους αισθητήρες.
Παρακολούθηση λειτουργιών
[επεξεργασία]Ένας άλλος τρόπος με τον οποίο αντιλαμβανόμαστε την κίνηση είναι μέσω της παρακολούθησης χαρακτηριστικών. Η Παρακολούθηση στοιχείων συνίσταται στην ανάλυση του εάν οι τοπικές λειτουργίες ενός αντικειμένου έχουν αλλάξει θέσεις ή όχι και υπονοούν τη μετακίνηση από αυτήν την αλλαγή. Σε αυτήν την ενότητα αναφέρονται ορισμένες λειτουργίες σχετικά με τους ιχνηλάτες λειτουργιών.
Οι ιχνηλάτες λειτουργιών αποτυγχάνουν όταν ένα κινούμενο ερέθισμα εμφανίζεται πολύ γρήγορα. Οι ιχνηλάτες λειτουργιών έχουν το πλεονέκτημα έναντι των αισθητήρων κίνησης ώστε να μπορούν να αντιλαμβάνονται την κίνηση ενός αντικειμένου, ακόμη και αν η κίνηση διαχωρίζεται από διαλείπον κενά διαστήματα. Μπορούν επίσης να διαχωρίσουν αυτά τα δύο στάδια (κινήσεις και κενά διαστήματα). Οι αισθητήρες κίνησης, από την άλλη πλευρά, απλώς θα ενσωμάτωναν τα κενά με το κινούμενο ερέθισμα και θα δουν μια συνεχή κίνηση. Οι ιχνηλάτες λειτουργιών λειτουργούν στις θέσεις των αναγνωρισμένων χαρακτηριστικών. Για το λόγο αυτό, έχουν ελάχιστο όριο απόστασης που ταιριάζει με την ακρίβεια με την οποία οι θέσεις των χαρακτηριστικών μπορούν να διακρίνονται. Οι ιχνηλάτες λειτουργιών δεν εμφανίζουν μετακινήσεις, οι οποίες είναι οπτικές ψευδαισθήσεις που προκαλούνται ως αποτέλεσμα οπτικής προσαρμογής. Οι συνέπειες της κίνησης εμφανίζονται όταν, μετά την παρατήρηση ενός κινούμενου ερεθίσματος, ένα σταθερό αντικείμενο φαίνεται να κινείται προς την αντίθετη κατεύθυνση του προηγουμένως παρατηρούμενου κινούμενου ερεθίσματος. Είναι αδύνατο για αυτόν τον μηχανισμό να παρακολουθεί πολλαπλές κινήσεις σε διαφορετικά μέρη του οπτικού πεδίου και ταυτόχρονα. Από την άλλη πλευρά, οι πολλαπλές κινήσεις δεν αποτελούν πρόβλημα για τους αισθητήρες κίνησης, επειδή λειτουργούν παράλληλα σε όλο το οπτικό πεδίο.
Πειράματα έχουν διεξαχθεί χρησιμοποιώντας τις παραπάνω πληροφορίες για να φτάσουν ενδιαφέροντα συμπεράσματα σχετικά με τους ιχνηλάτες χαρακτηριστικών. Τα πειράματα με σύντομα ερεθίσματα έχουν δείξει ότι τα σχέδια χρώματος και τα σχήματα αντίθεσης σε υψηλές αντιθέσεις δεν αντιλαμβάνονται οι ιχνηλάτες χαρακτηριστικών αλλά οι αισθητήρες κίνησης. Τα πειράματα με κενά διαστήματα έχουν επιβεβαιώσει ότι η παρακολούθηση χαρακτηριστικών μπορεί να συμβεί με κενά διαστήματα στην οθόνη. Μόνο σε μεγάλη αντίθεση, οι αισθητήρες κίνησης αντιλαμβάνονται την κίνηση των χρωματικών ερεθισμάτων και των μοντέλων αντίθεσης. Σε χαμηλές αντιθέσεις, οι ιχνηλάτες αναλύουν την κίνηση τόσο των χρωματογραφικών μοτίβων όσο και των φακέλων αντίθεσης και σε υψηλές αντιθέσεις οι αισθητήρες κίνησης αναλύουν τους φακέλους αντίθεσης. Τα πειράματα στα οποία τα άτομα κάνουν κρίσεις πολλαπλής κίνησης υποδηλώνουν ότι η παρακολούθηση χαρακτηριστικών είναι μια διαδικασία που πραγματοποιείται υπό συνειδητό έλεγχο και ότι είναι ο μόνος τρόπος που πρέπει να αναλύσουμε την κίνηση των φακέλων αντίθεσης σε οθόνες χαμηλής αντίθεσης. Αυτά τα αποτελέσματα συμφωνούν με την άποψη ότι η κίνηση των φακέλων αντίθεσης και των χρωμάτων εξαρτάται από την παρακολούθηση χαρακτηριστικών, εκτός εάν τα χρώματα είναι πολύ πάνω από το κατώφλι ή η μέση αντίθεση είναι υψηλή. Το κύριο συμπέρασμα αυτών των πειραμάτων είναι ότι πιθανότατα είναι η παρακολούθηση χαρακτηριστικών που επιτρέπει την αντίληψη των φακέλων αντίθεσης και των χρωμάτων.
Ψευδαισθήσεις κινήσεων
[επεξεργασία]Ως συνέπεια της διαδικασίας στην οποία λειτουργεί η ανίχνευση κίνησης, κάποιες στατικές εικόνες μπορεί να φανούν σαν να κινούνται. Αυτές οι εικόνες δίνουν μια εικόνα για τις υποθέσεις που κάνει το οπτικό σύστημα και ονομάζονται οπτικές ψευδαισθήσεις.
Μια διάσημη ψευδαίσθηση κινήσεων που σχετίζεται με σήματα κίνησης πρώτης τάξης είναι το φαινόμενο Phi, το οποίο είναι μια οπτική ψευδαίσθηση που μας κάνει να αντιλαμβανόμαστε την κίνηση αντί μιας ακολουθίας εικόνων. Αυτή η ψευδαίσθηση κινήσεων μας επιτρέπει να βλέπουμε ταινίες ως συνέχεια και όχι ως ξεχωριστές εικόνες. Το φαινόμενο phi επιτρέπει σε μια ομάδα παγωμένων εικόνων που αλλάζουν με σταθερή ταχύτητα να θεωρούνται ως μια συνεχής κίνηση. Το φαινόμενο Phi δεν πρέπει να συγχέεται με τη Βήτα Κίνηση, επειδή η πρώτη είναι μια εμφανής κίνηση που προκαλείται από φωτεινούς παλμούς σε μια ακολουθία, ενώ η τελευταία είναι μια εμφανής κίνηση που προκαλείται από φωτεινούς στατικές παλμούς.
Οι ψευδαισθήσεις κινήσεων συμβαίνουν όταν η αντίληψη κίνησης, η ανάλυση κίνησης και η ερμηνεία αυτών των σημάτων είναι παραπλανητικές και το οπτικό μας σύστημα δημιουργεί ψευδαισθήσεις σχετικά με την κίνηση. Αυτές οι ψευδαισθήσεις μπορούν να ταξινομηθούν ανάλογα με το ποια διαδικασία τους επιτρέπει να συμβούν. Οι ψευδαισθήσεις ταξινομούνται ως ψευδαισθήσεις που σχετίζονται με την ανίχνευση κίνησης, την ολοκλήρωση 2D και την τρισδιάστατη ερμηνεία.
Οι πιο δημοφιλείς ψευδαισθήσεις σχετικά με την ανίχνευση κίνησης είναι οι τετράχρονες κινήσεις, οι RDK και οι κινήσεις δεύτερης τάξης σηματοδοτούν ψευδαισθήσεις. Οι πιο δημοφιλείς ψευδαισθήσεις σχετικά με την ενσωμάτωση 2D είναι Motion Capture, Plaid Motion και Direct Repulsion. Ομοίως, αυτές που αφορούν την τρισδιάστατη ερμηνεία είναι η Μετασχηματιστική Κίνηση, το Κινητικό Βάθος, η Σκιώδης Κίνηση, η Βιολογική Κίνηση, η Στερεοκινητική κίνηση, η Κινητή Εικόνα και η κίνηση 2 Stroke. Υπάρχουν πολύ περισσότεροι κινούμενοι ψευδαισθήσεις, και όλα δείχνουν κάτι ενδιαφέρον για τους μηχανισμούς ανίχνευσης κινήσεων, ανίχνευσης κινήσεων και ανάλυσης. Για περισσότερες πληροφορίες, επισκεφθείτε τον ακόλουθο σύνδεσμο:
http://www.lifesci.sussex.ac.uk/home/George_Mather/Motion/
Άνοιγμα προβλημάτων
[επεξεργασία]Παρόλο που ακόμα δεν καταλαβαίνουμε τις περισσότερες λεπτομέρειες σχετικά με την αντίληψη κίνησης, η κατανόηση των μηχανισμών με τους οποίους γίνεται αντιληπτή η κίνηση καθώς και η ψευδαίσθηση της κίνησης μπορούν να δώσουν στον αναγνώστη μια καλή εικόνα της κατάστασης της τέχνης στο θέμα. Μερικά από τα ανοιχτά προβλήματα σχετικά με την αντίληψη κίνησης είναι οι μηχανισμοί σχηματισμού 3D εικόνων σε παγκόσμια κίνηση και το πρόβλημα διαφράγματος.
Παγκόσμια σήματα κίνησης από τον αμφιβληστροειδή είναι ενσωματωμένα για να φθάσουν σε ένα 2-διαστάσεων σήμα παγκόσμιας κίνησης? Ωστόσο, δεν είναι σαφές πώς διαμορφώνεται η 3D παγκόσμια κίνηση. Το πρόβλημα της διαφράγματος συμβαίνει επειδή κάθε πεδίο υποδοχής στο οπτικό σύστημα καλύπτει μόνο ένα μικρό κομμάτι του οπτικού κόσμου, το οποίο οδηγεί σε ασάφειες στην αντίληψη. Το πρόβλημα ανοίγματος αναφέρεται στο πρόβλημα ενός κινούμενου περιγράμματος το οποίο, όταν παρατηρείται τοπικά, είναι συνεπές με διαφορετικές δυνατότητες κίνησης. Αυτή η ασάφεια έχει γεωμετρική προέλευση - δεν μπορεί να ανιχνευθεί κίνηση παράλληλη με το περίγραμμα, καθώς οι αλλαγές σε αυτό το στοιχείο της κίνησης δεν αλλάζουν τις εικόνες που παρατηρούνται μέσω του ανοίγματος. Το μόνο συστατικό που μπορεί να μετρηθεί είναι η ταχύτητα ορθογώνια προς τον προσανατολισμό του περιγράμματος. Για το λόγο αυτό, η ταχύτητα του κινήματος θα μπορούσε να είναι οτιδήποτε από την οικογένεια κινήσεων κατά μήκος μιας γραμμής στον χώρο ταχύτητας. Αυτό το πρόβλημα ανοίγματος δεν παρατηρείται μόνο σε ευθεία περίγραμμα, αλλά και σε ομαλά καμπύλες, καθώς είναι κατά προσέγγιση ίσια όταν παρατηρείται τοπικά. Παρόλο που οι μηχανισμοί για την επίλυση του Προβλήματος Ανοιγμάτων είναι ακόμα άγνωστοι, υπάρχει κάποια υπόθεση για το πώς θα μπορούσε να λυθεί. Για παράδειγμα, θα μπορούσε να επιλυθεί αυτό το πρόβλημα συνδυάζοντας πληροφορίες σε ολόκληρο το διάστημα ή από διαφορετικά περιγράμματα του ίδιου αντικειμένου.
Συμπεράσματα
[επεξεργασία]Σε αυτό το κεφάλαιο, παρουσιάσαμε την αντίληψη κίνησης και τους μηχανισμούς με τους οποίους το οπτικό μας σύστημα εντοπίζει την κίνηση. Οι ψευδαισθήσεις κίνησης έδειξαν πώς τα σήματα κίνησης μπορεί να είναι παραπλανητικά και, συνεπώς, να οδηγούν σε εσφαλμένα συμπεράσματα σχετικά με την κίνηση. Είναι σημαντικό να θυμάστε ότι η αντίληψη κίνησης και η ανάλυση κίνησης δεν είναι οι ίδιες. Οι αισθητήρες κίνησης και οι ιχνηλάτες λειτουργιών αλληλοσυμπληρώνονται ώστε το οπτικό σύστημα να αντιλαμβάνεται την κίνηση.
Η αντίληψη κίνησης είναι περίπλοκη και εξακολουθεί να είναι μια ανοιχτή περιοχή έρευνας. Αυτό το κεφάλαιο περιγράφει μοντέλα σχετικά με τον τρόπο λειτουργίας των αισθητήρων κίνησης και υποθέσεις σχετικά με τα χαρακτηριστικά των ιχνηλατών χαρακτηριστικών. Ωστόσο, χρειάζονται περισσότερα πειράματα για να μάθουν για τα χαρακτηριστικά αυτών των μηχανισμών και να είναι σε θέση να κατασκευάσουν μοντέλα που μοιάζουν με τις πραγματικές διαδικασίες του οπτικού συστήματος με μεγαλύτερη ακρίβεια.
Η ποικιλία των μηχανισμών ανάλυσης κίνησης και αντίληψης κίνησης που περιγράφονται σε αυτό το κεφάλαιο, καθώς και η εκλέπτυνση των τεχνητών μοντέλων που έχουν σχεδιαστεί για να τα περιγράψουν, καταδεικνύουν ότι υπάρχει πολύ μεγάλη πολυπλοκότητα στον τρόπο με τον οποίο ο φλοιός επεξεργάζεται σήματα από το εξωτερικό περιβάλλον. Χιλιάδες εξειδικευμένοι νευρώνες ενσωματώνουν και ερμηνεύουν κομμάτια τοπικών σημάτων για να σχηματίσουν σφαιρικές εικόνες κινούμενων αντικειμένων στον εγκέφαλό μας. Η κατανόηση ότι πολλοί φορείς και διαδικασίες στο σώμα μας πρέπει να δουλεύουν μαζί για να αντιληφθούμε την κίνηση καθιστά την ικανότητά μας σε αυτό πιο αξιοσημείωτο ότι εμείς ως άνθρωποι μπορούμε να το κάνουμε με τέτοια ευκολία.
Bιβλιογραφικές Aναφορές
[επεξεργασία]Αντίληψη χρωμάτων
[επεξεργασία]Εισαγωγή
[επεξεργασία]Οι άνθρωποι (μαζί με τα πρωτεύοντα όπως οι μαϊμούδες και οι γορίλες) έχουν την καλύτερη αντίληψη χρώματος μεταξύ των θηλαστικών [1]. Ως εκ τούτου, δεν είναι τυχαίο ότι το χρώμα παίζει σημαντικό ρόλο σε μια ευρεία ποικιλία πτυχών. Για παράδειγμα, το χρώμα είναι χρήσιμο για τη διάκριση και την διαφοροποίηση αντικειμένων, επιφανειών, φυσικών τοπίων, ακόμη και προσώπων [2],[3]. Το χρώμα είναι επίσης ένα σημαντικό εργαλείο για τη μη λεκτική επικοινωνία, συμπεριλαμβανομένου του συναισθήματος [4].
Για πολλές δεκαετίες, ήταν μια πρόκληση να βρεθούν οι δεσμοί μεταξύ των φυσικών ιδιοτήτων του χρώματος και των αντιληπτικών ιδιοτήτων του. Συνήθως, αυτά μελετώνται με δύο διαφορετικές προσεγγίσεις: τη συμπεριφορική απόκριση που προκαλείται από το χρώμα (ονομάζεται επίσης ψυχοφυσική) και την πραγματική φυσιολογική απόκριση που προκαλείται από αυτήν [5].
Εδώ θα επικεντρωθούμε μόνο στην τελευταία. Η μελέτη της φυσιολογικής βάσης της έγχρωμης όρασης, για την οποία σχεδόν τίποτα δεν ήταν γνωστό πριν από το δεύτερο μισό του εικοστού αιώνα, έχει προχωρήσει αργά και σταθερά από το 1950. Σημαντική πρόοδος έχει σημειωθεί σε πολλούς τομείς, ειδικά σε επίπεδο υποδοχέα. Χάρη στις μεθόδους της μοριακής βιολογίας, ήταν δυνατό να αποκαλυφθούν άγνωστες λεπτομέρειες σχετικά με τη γενετική βάση των κηλίδων. Επιπλέον, όλο και περισσότερες περιοχές του φλοιού έχουν δειχθεί ότι επηρεάζονται από οπτικά ερεθίσματα, αν και η συσχέτιση της αντίληψης χρώματος με τη φυσιολογική δραστηριότητα που εξαρτάται από το μήκος κύματος πέρα από τους υποδοχείς δεν είναι τόσο εύκολο να διακριθεί [6].
Σε αυτό το κεφάλαιο, στοχεύουμε να εξηγήσουμε τα βασικά στοιχεία των διαφορετικών διεργασιών της αντίληψης χρώματος κατά μήκος της οπτικής διαδρομής, από τον αμφιβληστροειδή στο μάτι έως τον οπτικό φλοιό στον εγκέφαλο. Για ανατομικές λεπτομέρειες, ανατρέξτε στο Κεφ. "Ανατομία του οπτικού συστήματος" αυτού του Wikibook.
Χρωματική αντίληψη στη αμφιβληστροειδή
[επεξεργασία]Όλα τα χρώματα που μπορούν να διακριθούν από τον άνθρωπο μπορούν να παραχθούν από το μίγμα μόνο τριών βασικών (βασικών) χρωμάτων. Εμπνευσμένο από αυτή την ιδέα της ανάμειξης χρωμάτων, έχει προταθεί ότι το χρώμα υποστηρίζεται από τρεις κατηγορίες αισθητήρων, καθεμία από τις οποίες έχει μέγιστη ευαισθησία σε διαφορετικό τμήμα του ορατού φάσματος [1]. Αρχικά προτάθηκε ρητά το 1853 ότι υπάρχουν τρεις βαθμοί ελευθερίας στη φυσιολογική αντιστοίχιση χρωμάτων [7]. Αυτό επιβεβαιώθηκε αργότερα το 1886 [8] (με αξιοσημείωτα στενά αποτελέσματα σε πρόσφατες μελέτες [9], [10]). Αυτοί οι προτεινόμενοι αισθητήρες χρώματος είναι στην πραγματικότητα οι λεγόμενοι κώνοι (Σημείωση: Σε αυτό το κεφάλαιο, θα ασχοληθούμε μόνο με τους κώνους.) Οι ραβδία συμβάλλουν στην όραση μόνο σε χαμηλά επίπεδα φωτός Αν και είναι γνωστό ότι επηρεάζουν την αντίληψη των χρωμάτων, μικρές και μπορούν να αγνοηθούν εδώ.) [11]. Τα κώνοι είναι από τους δύο τύπους κυττάρων φωτοϋποδοχέα που βρίσκονται στον αμφιβληστροειδή, με σημαντική συγκέντρωση αυτών στον φούντα. Ο παρακάτω πίνακας περιλαμβάνει τους τρεις τύπους κελιών κώνου. Αυτά διακρίνονται από διαφορετικούς τύπους χρωστικής ροδοψίνης. Οι αντίστοιχες καμπύλες απορρόφησης φαίνονται στο παρακάτω σχήμα.
| Name | Higher sensitivity to color | Absorption curve peak [nm] |
|---|---|---|
| S, SWS, B | Blue | 420 |
| M, MWS, G | Green | 530 |
| L, LWS, R | Red | 560 |
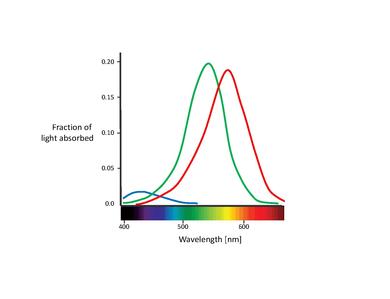
Αν και δεν έχει επιτευχθεί συναίνεση για την ονομασία των διαφορετικών τύπων κώνου, οι πιο ευρέως χρησιμοποιούμενες ονομασίες αναφέρονται είτε στην κορυφή του φάσματος δράσης είτε στο χρώμα στο οποίο είναι ευαίσθητοι (κόκκινο, πράσινο, μπλε) [6]. Σε αυτό το κείμενο, θα χρησιμοποιήσουμε τον ορισμό S-M-L (για βραχύ, μεσαίο και μακρύ μήκος κύματος), αφού αυτά τα ονόματα είναι καταλληλότερα περιγραφικά. Η μπλε-πράσινη-κόκκινη ονοματολογία είναι κάπως παραπλανητική, αφού όλοι οι τύποι κώνων είναι ευαίσθητοι σε ένα μεγάλο εύρος μηκών κύματος.
Ένα σημαντικό χαρακτηριστικό σχετικά με τους τρεις τύπους κώνου είναι η σχετική τους κατανομή στον αμφιβληστροειδή. Αποδεικνύεται ότι οι S-κώνες παρουσιάζουν σχετικά χαμηλή συγκέντρωση μέσω του αμφιβληστροειδούς, λείπουν εντελώς στην πιο κεντρική περιοχή του φούντα. Στην πραγματικότητα, είναι πολύ ευρύχωρα ώστε να διαδραματίζουν σημαντικό ρόλο στην χωρική όραση, παρόλο που είναι ικανά να διαμεσολαβούν στην αδύναμη συνοριακή αντίληψη [12]. Η φόβα κυριαρχείται από L- και M-κώνους. Η αναλογία των δύο τελευταίων μετριέται συνήθως ως λόγος. Έχουν αναφερθεί διαφορετικές τιμές για τον λόγο L / M, που κυμαίνονται από 0,67 [13] έως 2 [14], με τις τελευταίες να είναι οι πλέον αποδεκτές. Γιατί οι L-cones σχεδόν πάντα ξεπερνούν τους M-κώνους παραμένει ασαφής. Παραδόξως, ο λόγος σχετικού κώνου δεν έχει σχεδόν καμία σημαντική επίδραση στην έγχρωμη όραση. Αυτό δείχνει σαφώς ότι ο εγκέφαλος είναι πλαστικός, ικανός να αποκτήσει νόημα από οποιαδήποτε σήματα κώνου λαμβάνει [15], [16].
Είναι επίσης σημαντικό να σημειωθεί η αλληλεπικάλυψη των φασμάτων απορρόφησης L- και M-κώνου. Ενώ το φάσμα απορρόφησης S-κώνου διαχωρίζεται σαφώς, οι κορυφές L- και M-κώνου απέχουν μόνο περίπου 30 nm, ενώ οι φασματικές καμπύλες τους αλληλεπικαλύπτονται επίσης σημαντικά. Αυτό έχει ως αποτέλεσμα μια υψηλή συσχέτιση στα αλιεύματα φωτονίων αυτών των δύο κατηγοριών κώνου. Αυτό εξηγείται από το γεγονός ότι για να επιτευχθεί η μέγιστη δυνατή οξύτητα στο κέντρο της φούντας, το οπτικό σύστημα αντιμετωπίζει τους κώνους L και M εξίσου χωρίς να λαμβάνει υπόψη τα φάσματα απορρόφησης τους. Επομένως, κάθε είδους διαφορά οδηγεί σε υποβάθμιση του σήματος φωτεινότητας [17]. Με άλλα λόγια, ο μικρός διαχωρισμός μεταξύ των φασμάτων L- και M-κώνων μπορεί να ερμηνευθεί ως συμβιβασμός μεταξύ των αναγκών για έγχρωμη όραση υψηλής αντίθεσης και όραση φωτεινότητας υψηλής ακρίβειας. Αυτό είναι σύμφωνο με την έλλειψη S-κώνων στο κεντρικό τμήμα του fovea, όπου η οπτική οξύτητα είναι υψηλότερη. Επιπλέον, η στενή απόσταση των φασμάτων απορρόφησης L- και M-κώνων μπορεί επίσης να εξηγηθεί από τη γενετική τους προέλευση. Και οι δύο τύποι κώνων υποτίθεται ότι έχουν εξελιχθεί "πρόσφατα" (περίπου 35 εκατομμύρια χρόνια πριν) από έναν κοινό πρόγονο, ενώ οι S-κώνοι πιθανώς αποχωρίστηκαν από τον προγονικό παλμό πολύ νωρίτερα [11].
Οι λειτουργίες φασματικής απορρόφησης των τριών διαφορετικών τύπων κωνικών κυττάρων είναι το χαρακτηριστικό γνώρισμα της ανθρώπινης έγχρωμης όρασης. Αυτή η θεωρία επιλύει ένα πολύ γνωστό πρόβλημα: παρόλο που μπορούμε να δούμε εκατομμύρια διαφορετικά χρώματα (οι άνθρωποι μπορούν να διακρίνουν 7 έως 10 εκατομμύρια διαφορετικά χρώματα, οι αμφιβληστροειδοί απλά δεν έχουν αρκετό χώρο για να χωρέσουν μεμονωμένους ανιχνευτές για κάθε χρώμα σε κάθε θέση αμφιβληστροειδούς.
Από τη Ρετίνα στον Εγκέφαλο
[επεξεργασία]Τα σήματα που μεταδίδονται από τον αμφιβληστροειδή σε υψηλότερα επίπεδα δεν είναι απλές σημειακές αναπαραστάσεις των σημάτων υποδοχέα, αλλά μάλλον αποτελούνται από εξελιγμένους συνδυασμούς των σημάτων υποδοχέα. Ο στόχος αυτής της ενότητας είναι να παράσχει μια σύντομη περιγραφή των διαδρομών που λαμβάνουν ορισμένες από αυτές τις πληροφορίες.
Αφού η οπτική εικόνα στον αμφιβληστροειδή μεταδιεγείρεται σε χημικά και ηλεκτρικά σήματα στους φωτοϋποδοχείς, τα σήματα με διαμόρφωση πλάτους μετατρέπονται σε παραστάσεις διαμορφωμένες σε συχνότητα στο κυψελιδικό κύτταρο και σε υψηλότερα επίπεδα. Σε αυτά τα νευρικά κύτταρα, το μέγεθος του σήματος αντιπροσωπεύεται από τον αριθμό των τάσεων τάσης ανά δευτερόλεπτο που τροφοδοτούνται από το κύτταρο και όχι από τη διαφορά τάσης κατά μήκος της κυτταρικής μεμβράνης. Προκειμένου να εξηγηθούν και να αναπαρασταθούν οι φυσιολογικές ιδιότητες αυτών των κυττάρων, θα βρούμε πολύ χρήσιμη την έννοια των δεκτικών πεδίων.
Ένα δεκτικό πεδίο είναι μια γραφική αναπαράσταση της περιοχής στο οπτικό πεδίο στο οποίο αποκρίνεται ένα δεδομένο κύτταρο. Επιπλέον, η φύση της απόκρισης τυπικά υποδεικνύεται για διάφορες περιοχές του δεκτικού πεδίου. Για παράδειγμα, μπορούμε να θεωρήσουμε το πεδίο υποδοχής ενός φωτοϋποδοχέα ως μια μικρή κυκλική περιοχή που αντιπροσωπεύει το μέγεθος και τη θέση της ευαισθησίας αυτού του συγκεκριμένου υποδοχέα στο οπτικό πεδίο. Το σχήμα που ακολουθεί δείχνει παραδειγματικά πεδία υποδοχής για γαγγλιακά κύτταρα, τυπικά σε ανταγωνισμό με κέντρο-περιβάλλον. Το αριστερό πεδίο υποδοχής στο σχήμα απεικονίζει μια θετική κεντρική απόκριση (γνωστή ως επί τόπου). Αυτό το είδος απόκρισης παράγεται συνήθως από μια θετική εισροή από ένα μόνο κώνο που περιβάλλεται από αρνητική απόκριση που παράγεται από πολλούς γειτονικούς κώνους. Επομένως, η απόκριση αυτού του κυττάρου γαγγλίου θα αποτελείται από εισροές από διάφορους κώνους με θετικά και αρνητικά σημεία. Με αυτό τον τρόπο, η κυψέλη δεν ανταποκρίνεται μόνο σε σημεία φωτός, αλλά χρησιμεύει ως ανιχνευτής άκρης (ή πιο σωστά, σημείο). Κατ 'αναλογία με την ορολογία των υπολογιστικών οραμάτων, μπορούμε να σκεφτούμε τις αποκρίσεις κυττάρων γαγγλίου ως αποτέλεσμα μιας συνέλιξης με έναν πυρήνα ανιχνευτή άκρων. Το δεξιό πεδίο υποδοχής στο σχήμα απεικονίζει μια αρνητική κεντρική απόκριση (γνωστή ως off-center), η οποία είναι εξίσου πιθανή. Συνήθως, τα κύτταρα στο κέντρο και εκτός κέντρου θα εμφανιστούν στην ίδια χωρική θέση, τροφοδοτούμενα από τους ίδιους φωτοϋποδοχείς, με αποτέλεσμα μια ενισχυμένη δυναμική εμβέλεια.
Το κατώτερο σχήμα δείχνει ότι εκτός από τον χωρικό ανταγωνισμό, τα γαγγλιακά κύτταρα μπορούν επίσης να έχουν φασματική αντιπαράθεση. Για παράδειγμα, το αριστερό μέρος του κατώτερου σχήματος απεικονίζει μια κόκκινη-πράσινη ανταπόκριση του αντιπάλου με το κέντρο που τροφοδοτείται με θετική είσοδο από έναν L-κώνο και το περιβάλλον που τροφοδοτείται από μια αρνητική είσοδο από Μ-κώνους. Από την άλλη πλευρά, το δεξιό μέρος του κατώτερου σχήματος απεικονίζει την εκτός κέντρου έκδοση αυτής της κελιά. Ως εκ τούτου, προτού οι οπτικές πληροφορίες έχουν βγει από τον αμφιβληστροειδή, έχει ήδη συμβεί επεξεργασία, με βαθιά επίδραση στην εμφάνιση χρώματος. Υπάρχουν και άλλοι τύποι και ποικιλίες απαντήσεων κυττάρων γαγγλίου, αλλά όλες μοιράζονται αυτές τις βασικές έννοιες.
Στο δρόμο προς τον πρωτογενή οπτικό φλοιό, οι άξονες των γαγγλιακών κυττάρων συγκεντρώνονται για να σχηματίσουν το οπτικό νεύρο, το οποίο προβάλλει στον πλευρικό πυρήνα γονιδιώματος (LGN) στον θάλαμο. Η κωδικοποίηση στο οπτικό νεύρο είναι εξαιρετικά αποτελεσματική, διατηρώντας τον αριθμό των νευρικών ινών στο ελάχιστο (περιοριζόμενο από το μέγεθος του οπτικού νεύρου) και έτσι το μέγεθος του τυφλού σημείου του αμφιβληστροειδούς όσο το δυνατόν μικρότερο (περίπου 5 ° πλάτος κατά 7 ° υψηλός). Επιπλέον, τα παρουσιαζόμενα κύτταρα γαγγλίων δεν θα έχουν καμία απάντηση σε ομοιόμορφο φωτισμό, καθώς οι θετικές και οι αρνητικές περιοχές είναι ισορροπημένες. Με άλλα λόγια, τα μεταδιδόμενα σήματα δεν είναι συνδεδεμένα. Για παράδειγμα, οι πληροφορίες από τα γειτονικά τμήματα των φυσικών σκηνών είναι πολύ συσχετισμένες χωρικά και συνεπώς εξαιρετικά προβλέψιμες [18]. Η πλευρική αναστολή μεταξύ γειτονικών γαγγλιοκυττάρων αμφιβληστροειδούς ελαχιστοποιεί αυτή τη χωρική συσχέτιση, βελτιώνοντας έτσι την αποτελεσματικότητα. Μπορούμε να το δούμε ως διαδικασία συμπίεσης εικόνων που πραγματοποιείται στον αμφιβληστροειδή χιτώνα.
Δεδομένης της αλληλεπικάλυψης των φασμάτων απορρόφησης L- και M-κώνου, τα σήματα τους είναι επίσης πολύ συσχετισμένα. Σε αυτή την περίπτωση, η αποτελεσματικότητα της κωδικοποίησης βελτιώνεται συνδυάζοντας τα σήματα κώνου προκειμένου να ελαχιστοποιηθεί η εν λόγω συσχέτιση. Μπορούμε να το κατανοήσουμε πιο εύκολα χρησιμοποιώντας την κύρια ανάλυση στοιχείων (PCA). Το PCA είναι μια στατιστική μέθοδος που χρησιμοποιείται για να μειώσει τη διαστατικότητα ενός δεδομένου συνόλου μεταβλητών μετασχηματίζοντας τις αρχικές μεταβλητές σε ένα σύνολο νέων μεταβλητών, τα κύρια συστατικά (PCs). Ο πρώτος υπολογιστής αντιπροσωπεύει ένα μέγιστο ποσό συνολικής διακύμανσης στις αρχικές μεταβλητές, ο δεύτερος υπολογιστής αντιστοιχεί σε ένα μέγιστο ποσό διακύμανσης που δεν υπολογίστηκε από το πρώτο στοιχείο και ούτω καθεξής. Επιπλέον, οι υπολογιστές είναι γραμμικώς ανεξάρτητοι και ορθογώνιοι ο ένας στον άλλο στο χώρο των παραμέτρων. Το βασικό πλεονέκτημα της PCA είναι ότι μόνο μερικοί από τους ισχυρότερους υπολογιστές είναι αρκετοί για να καλύψουν τη συντριπτική πλειοψηφία της μεταβλητότητας του συστήματος [19]. Αυτό το σχήμα έχει χρησιμοποιηθεί με τις λειτουργίες απορρόφησης κώνου [20] και ακόμη και με τα φυσικώς απαντώμενα φάσματα [21],[22]. Οι υπολογιστές που βρέθηκαν στο χώρο των κωνικών διεγέρσεων που παράγονται από φυσικά αντικείμενα είναι 1) ένας άξονας φωτεινότητας όπου προστίθενται τα σήματα L- και M-κώνων (L + M), 2) η διαφορά του L- και Mκώνου (LM), και 3) έναν άξονα χρώματος όπου το σήμα S-κώνου διαφοροποιείται με το άθροισμα των σημάτων L- και M-κώνων (S- (L + M)). Αυτά τα κανάλια, που προέρχονται από μια μαθηματική / υπολογιστική προσέγγιση, συμπίπτουν με τους τρεις δίαυλους αμφιβληστροειδούς που ανευρίσκονται σε ηλεκτροφυσιολογικά πειράματα [23],[24]. Χρησιμοποιώντας αυτούς τους μηχανισμούς, οι οπτικές περιττές πληροφορίες εξαλείφονται στον αμφιβληστροειδή χιτώνα.
Υπάρχουν τρία κανάλια πληροφοριών που στην πραγματικότητα επικοινωνούν αυτές τις πληροφορίες από τον αμφιβληστροειδή μέσω των κυττάρων των γαγγλίων στο LGN. Διαφέρουν όχι μόνο στις χρωματικές ιδιότητές τους, αλλά και στο ανατομικό τους υπόστρωμα. Αυτά τα κανάλια δημιουργούν σημαντικούς περιορισμούς για βασικές εργασίες χρωμάτων, όπως ανίχνευση και διάκριση.
Στο πρώτο κανάλι, η έξοδος των L- και M-κώνων μεταδίδεται συνεργιστικά σε διάχυτα διπολικά κύτταρα και στη συνέχεια σε κύτταρα στα μαγνητικά στρώματα (Μ-) του LGN (δεν πρέπει να συγχέεται με τους Μ-κώνους του αμφιβληστροειδούς) [25]. Τα δεκτικά πεδία των Μ-κυττάρων αποτελούνται από ένα κέντρο και ένα περιβάλλον, τα οποία είναι χωρικά ανταγωνιστικά. Τα Μκύτταρα έχουν ευαισθησία υψηλής αντίθεσης για τα ερεθίσματα φωταύγειας, αλλά δεν δείχνουν απόκριση σε κάποιο συνδυασμό των εισροών του αντιπάλου LM [26].
Ωστόσο, επειδή τα μηδενικά σημεία διαφορετικών κυττάρων Μ ποικίλλουν ελαφρώς, η αντίδραση του πληθυσμού δεν είναι ποτέ πραγματικά μηδενική. Αυτή η ιδιότητα μεταφέρεται πραγματικά σε φλοιώδεις περιοχές με κυρίαρχη είσοδο κυψελών Μ [26].
Η μονοκυτταρική οδός (Ρ-) προέρχεται από τις μεμονωμένες εξόδους από L- ή Μ-κώνο σε διπολικά κύτταρα του μαστού. Αυτές παρέχουν είσοδο σε Ρ-κύτταρα του αμφιβληστροειδούς [11]. Στον φούντα, τα κέντρα δεκτικών πεδίων των κυττάρων Ρ σχηματίζονται από απλούς L- ή Μ-κώνους. Η δομή του περιβάλλουμενου πεδίου του P-κυττάρου εξακολουθεί να συζητείται. Ωστόσο, η πιο αποδεκτή θεωρία δηλώνει ότι το περίγραμμα αποτελείται από έναν ειδικό τύπο κώνου, καταλήγοντας σε ένα πεδίο υποδοχής του χώρου αντιπάλου για ερεθίσματα φωταύγειας [27]. Τα μεσοκυτταρικά στρώματα συμβάλλουν με το 80% περίπου των συνολικών προβολών από τον αμφιβληστροειδή στο LGN [28].
Τέλος, η πρόσφατα ανακαλυφθείσα οδοκυτταρική οδός (Κ-) μεταφέρει κυρίως σήματα από S-κώνους [29]. Ομάδες αυτού του τύπου κώνων προβάλλουν ειδικά διπολικά κύτταρα, τα οποία με τη σειρά τους παρέχουν εισροή σε συγκεκριμένα μικρά γαγγλιακά κύτταρα. Αυτά συνήθως δεν είναι χωρικά αντίπαλοι. Οι άξονες των μικρών γαγγλιοκυττάρων προβάλλουν σε λεπτά στρώματα του LGN (δίπλα στα πρωτογενή στρώματα) [30].
Ενώ τα κύτταρα των γαγγλιοκυττάρων καταλήγουν στο LGN (δημιουργώντας συνάψεις με κύτταρα LGN), φαίνεται να υπάρχει μία αντιστοιχία μεταξύ των γαγγλιοκυττάρων και των κυττάρων LGN. Το LGN φαίνεται να λειτουργεί ως σταθμός αναμετάδοσης για τα σήματα. Ωστόσο, εξυπηρετεί πιθανώς κάποια οπτική λειτουργία, καθώς υπάρχουν νευρικές προβολές από τον φλοιό πίσω στο LGN που θα μπορούσαν να χρησιμεύσουν ως κάποιο είδος μηχανισμού ανάδρασης ή προσαρμογής. Οι άξονες των κυττάρων LGN προβάλλουν στην οπτική περιοχή ένα (V1) στον οπτικό φλοιό στον ινιακό λοβό.
Χρωματική αντίληψη στον εγκέφαλο
[επεξεργασία]Στον φλοιό, οι προεξοχές από τις μαγνητικές, παρβο- και κυνοκυτταρικές οδούς καταλήγουν σε διαφορετικά στρώματα του πρωτογενούς οπτικού φλοιού. Οι μαγνητοκυτταρικές ίνες νευρώνουν κυρίως το στρώμα 4Cα και το στρώμα 6. Οι παραβολοειδείς νευρώνες προεξέχουν ως επί το πλείστον σε 4Cβ και στα στρώματα 4Α και 6. Οι κωνοκυτταρικοί νευρώνες τερματίζουν στις πλούσιες σε κυτόχρωμα οξειδάσες (CO) στα στρώματα 1, 2 και 3 [31].
Μόλις βρεθεί στον οπτικό φλοιό, η κωδικοποίηση των οπτικών πληροφοριών γίνεται σημαντικά πιο περίπλοκη. Με τον ίδιο τρόπο τα αποτελέσματα των διαφόρων φωτοϋποδοχέων συνδυάζονται και συγκρίνονται με την παραγωγή αποκρίσεων κυττάρων γαγγλίου, τα αποτελέσματα διαφόρων κυττάρων LGN συγκρίνονται και συνδυάζονται για την παραγωγή φλοιώδους αποκρίσεως. Καθώς τα σήματα προχωρούν περισσότερο στην αλυσίδα επεξεργασίας των φλοιών, αυτή η διαδικασία επαναλαμβάνεται με ένα ταχέως αυξανόμενο επίπεδο πολυπλοκότητας μέχρι το σημείο που τα δεκτικά πεδία αρχίζουν να χάνουν νόημα. Ωστόσο, ορισμένες λειτουργίες και διαδικασίες έχουν εντοπιστεί και μελετηθεί σε συγκεκριμένες περιοχές του οπτικού φλοιού.
Στην περιοχή V1 (πτερυγικός φλοιός), οι διπλοί αντίπαλοι νευρώνες - οι νευρώνες που έχουν τα δεκτικοί τους τομείς τόσο χρωματικά όσο και χωρικά αντίθετοι σε σχέση με τις περιοχές on / off ενός μόνο δεκτικού πεδίου - συγκρίνουν τα έγχρωμα σήματα στον οπτικό χώρο [32]. Αποτελούν μεταξύ 5 και 10% των κυττάρων στο V1. Το χονδροειδές μέγεθος και το μικρό τους ποσοστό ταιριάζουν με την κακή χωρική ανάλυση της έγχρωμης όρασης [1]. Επιπλέον, δεν είναι ευαίσθητα στην κατεύθυνση των κινούμενων ερεθισμάτων (σε αντίθεση με κάποιους άλλους νευρώνες V1) και, ως εκ τούτου, είναι απίθανο να συνεισφέρουν στην αντίληψη κίνησης [33]. Ωστόσο, δεδομένης της εξειδικευμένης υποδοχής πεδίου δομή τους, αυτά τα είδη των κυττάρων είναι η νευρωνική βάση για τα έγχρωμα αποτελέσματα αντίθεσης, καθώς και ένα αποτελεσματικό μέσο για την κωδικοποίηση του ίδιου του χρώματος [34], [35]. Άλλα κύτταρα V1 ανταποκρίνονται σε άλλους τύπους ερεθισμάτων, όπως προσανατολισμένες ακμές, διάφορες χωρικές και χρονικές συχνότητες, συγκεκριμένες χωρικές τοποθεσίες και συνδυασμοί αυτών των χαρακτηριστικών, μεταξύ άλλων. Επιπλέον, μπορούμε να βρούμε κύτταρα που συνδυάζουν γραμμικά εισροές από κύτταρα LGN καθώς και κύτταρα που εκτελούν μη γραμμικό συνδυασμό. Αυτές οι απαντήσεις είναι απαραίτητες για την υποστήριξη προηγμένων οπτικών δυνατοτήτων, όπως το ίδιο το χρώμα.

Υπάρχουν σημαντικά λιγότερες πληροφορίες σχετικά με τις χρωματικές ιδιότητες των απλών νευρώνων στο V2 σε σύγκριση με το V1. Από μια πρώτη ματιά, φαίνεται ότι δεν υπάρχουν σημαντικές διαφορές στην κωδικοποίηση των χρωμάτων στα V1 και V2 [36]. Μια εξαίρεση από αυτό είναι η εμφάνιση μιας νέας τάξης κυτταρικού χρώματος κυττάρου [37]. Επομένως, έχει προταθεί ότι η περιοχή V2 συμμετέχει στην επεξεργασία της απόχρωσης. Ωστόσο, αυτό εξακολουθεί να είναι πολύ αμφιλεγόμενο και δεν έχει επιβεβαιωθεί.
Ακολουθώντας την αρθρωτή έννοια που αναπτύχθηκε μετά την ανακάλυψη της λειτουργικής οφθαλμολογικής δεσπόζουσας θέσης στο V1 και λαμβάνοντας υπόψη τον ανατομικό διαχωρισμό μεταξύ των οδών P, M και K (που περιγράφονται στο κεφάλαιο 3), προτάθηκε ότι ένα εξειδικευμένο σύστημα μέσα στο οπτικό πρέπει να υπάρχει ο φλοιός που είναι αφιερωμένος στην ανάλυση των πληροφοριών χρώματος [38]. Το V4 είναι η περιοχή που έχει προσελκύσει ιστορικά την περισσότερη προσοχή ως την πιθανή "περιοχή χρώματος" του εγκεφάλου. Αυτό συμβαίνει λόγω μιας επιρροής μελέτης που ισχυρίστηκε ότι το V4 περιείχε 100% των εκλεκτικών χρωστικών [39]. Ωστόσο, ο ισχυρισμός αυτός αμφισβητήθηκε από αρκετές μετέπειτα μελέτες, μερικοί από τους οποίους ανέφεραν ότι μόνο το 16% των νευρώνων V4 παρουσιάζουν συντονισμό απόχρωσης [40]. Επί του παρόντος, η πιο αποδεκτή ιδέα είναι ότι το V4 συμβάλλει όχι μόνο στο χρώμα, αλλά και στη διαμόρφωση της αντίληψης, της οπτικής προσοχής και της στερεοτυπίας. Επιπλέον, πρόσφατες μελέτες έχουν επικεντρωθεί σε άλλες περιοχές του εγκεφάλου που προσπαθούν να βρουν την "περιοχή χρωμάτων" του εγκεφάλου, όπως το TEO [41] και το PITd [42]. Η σχέση αυτών των περιφερειών μεταξύ τους εξακολουθεί να συζητείται. Για να συμβιβαστεί η συζήτηση, κάποιοι χρησιμοποιούν τον όρο οπίσθιο κατώτερο χρονικό (PIT) φλοιό για να υποδηλώσουν την περιοχή που περιλαμβάνει τα V4, TEO και PITd [1].
Εάν η φλοιώδης απόκριση στα κύτταρα V1, V2 και V4 είναι ήδη πολύ περίπλοκη, το επίπεδο πολυπλοκότητας των σύνθετων οπτικών αποκρίσεων σε ένα δίκτυο περίπου 30 οπτικών ζωνών είναι τεράστιο. Το σχήμα 4 δείχνει ένα μικρό τμήμα της συνδεσιμότητας των διαφορετικών φλοιωδών περιοχών (όχι των κυττάρων) που έχουν ταυτοποιηθεί [43].
Σε αυτό το στάδιο, γίνεται εξαιρετικά δύσκολο να εξηγηθεί η λειτουργία των κυττάρων μεμονωμένων φλοιών με απλούς όρους. Στην πραγματικότητα, η λειτουργία ενός μόνο κυττάρου μπορεί να μην έχει νόημα, αφού η αναπαράσταση διαφόρων αντιλήψεων πρέπει να κατανέμεται σε συλλογές κυττάρων σε ολόκληρο τον φλοιό.
Μηχανισμοί προσαρμογής χρωμάτων
[επεξεργασία]Αν και οι ερευνητές προσπαθούν να εξηγήσουν την επεξεργασία των σημάτων χρώματος στο ανθρώπινο οπτικό σύστημα, είναι σημαντικό να καταλάβουμε ότι η αντίληψη χρώματος δεν είναι μια σταθερή διαδικασία. Στην πραγματικότητα, υπάρχουν διάφοροι δυναμικοί μηχανισμοί που χρησιμεύουν για τη βελτιστοποίηση της οπτικής απόκρισης ανάλογα με το περιβάλλον προβολής. Ιδιαίτερη σημασία για την αντίληψη χρώματος είναι οι μηχανισμοί σκοτεινής, ελαφριάς και χρωματικής προσαρμογής.
Σκοτεινή προσαρμογή
[επεξεργασία]Η σκοτεινή προσαρμογή αναφέρεται στην αλλαγή στην οπτική ευαισθησία που συμβαίνει όταν μειώνεται το επίπεδο φωτισμού. Η απόκριση του οπτικού συστήματος σε μειωμένο φωτισμό είναι να γίνει πιο ευαίσθητη, αυξάνοντας την ικανότητά του να παράγει μια ουσιαστική οπτική απόκριση ακόμη και όταν οι συνθήκες φωτισμού δεν είναι βέλτιστες [44].
Το σχήμα 5 δείχνει την ανάκτηση της οπτικής ευαισθησίας μετά τη μετάβαση από ένα εξαιρετικά υψηλό επίπεδο φωτισμού σε ένα πλήρες σκοτάδι [43]. Κατ 'αρχάς, οι κώνοι γίνονται σταδιακά πιο ευαίσθητοι, έως ότου η καμπύλη ξεπεράσει μετά από μερικά λεπτά. Στη συνέχεια, μετά από περίπου 10 λεπτά, η οπτική ευαισθησία είναι σχεδόν σταθερή. Σε αυτό το σημείο, το σύστημα ράβδων, με μεγαλύτερο χρόνο ανάκτησης, έχει ανακτήσει αρκετή ευαισθησία ώστε να ξεπεράσει τους κώνους και επομένως να ανακτήσει τον έλεγχο της συνολικής ευαισθησίας. Η ευαισθησία της ραβδίου βελτιώνεται σταδιακά, μέχρι να γίνει ασυμπτωτική μετά από περίπου 30 λεπτά. Με άλλα λόγια, οι κώνοι είναι υπεύθυνοι για την ανάκτηση ευαισθησίας για τα πρώτα 10 λεπτά. Στη συνέχεια, οι ραβδία ξεπερνούν τους κώνους και αποκτούν πλήρη ευαισθησία μετά από περίπου 30 λεπτά.
Αυτός είναι μόνο ένας από τους πολλούς νευρικούς μηχανισμούς που παράγονται προκειμένου να προσαρμόζονται στις συνθήκες σκοτεινής κεραίας όσο το δυνατόν καλύτερο. Κάποιοι άλλοι νευρωνικοί μηχανισμοί περιλαμβάνουν το γνωστό αντανακλαστικό των μαθητών, την εξάντληση και την αναγέννηση της φωτοχρωματογραφίας, τον έλεγχο κέρδους στα κύτταρα του αμφιβληστροειδούς και άλλους μηχανισμούς υψηλότερου επιπέδου και τη γνωστική ερμηνεία μεταξύ άλλων.
Προσαρμογή φωτός
[επεξεργασία]Η προσαρμογή φωτός είναι ουσιαστικά η αντίστροφη διαδικασία της σκοτεινής προσαρμογής. Στην πραγματικότητα, οι βασικοί φυσιολογικοί μηχανισμοί είναι οι ίδιοι και για τις δύο διαδικασίες. Ωστόσο, είναι σημαντικό να το εξετάσουμε χωριστά, καθώς οι οπτικές του ιδιότητες διαφέρουν.

Η προσαρμογή φωτός συμβαίνει όταν αυξάνεται το επίπεδο φωτισμού. Επομένως, το οπτικό σύστημα πρέπει να γίνει λιγότερο ευαίσθητο για να παράγει χρήσιμες αντιλήψεις, δεδομένου ότι υπάρχει πολύ πιο ορατό φως διαθέσιμο. Το οπτικό σύστημα διαθέτει μια περιορισμένη δυναμική περιοχή εξόδου διαθέσιμη για τα σήματα που παράγουν τις αντιλήψεις μας. Ωστόσο, ο πραγματικός κόσμος έχει επίπεδα φωτισμού που καλύπτουν τουλάχιστον 10 τάξεις μεγέθους περισσότερο. Ευτυχώς, σπάνια πρέπει να βλέπουμε όλο το φάσμα των επιπέδων φωτισμού ταυτόχρονα.
Σε υψηλά επίπεδα φωτισμού, η προσαρμογή επιτυγχάνεται με λεύκανση με φωτοχρωστικές ουσίες. Αυτό κλιμακώνει τη δέσμευση φωτονίων στους υποδοχείς και προστατεύει την απόκριση του κώνου από κορεσμό σε φωτεινά υπόβαθρα. Οι μηχανισμοί προσαρμογής φωτός εμφανίζονται κυρίως στον αμφιβληστροειδή [45]. Στην πραγματικότητα, οι μεταβολές κέρδους είναι σε μεγάλο βαθμό κωνικές και τα σήματα προσαρμογής συγκεντρώνονται σε περιοχές που δεν είναι μεγαλύτερες από τη διάμετρο των μεμονωμένων κώνων [46], [47]. Αυτό δείχνει τον εντοπισμό προσαρμογής φωτός που μπορεί να είναι ήδη από τους υποδοχείς. Ωστόσο, φαίνεται ότι υπάρχουν περισσότερες από μία τοποθεσίες κλιμάκωσης ευαισθησίας. Ορισμένες από τις αλλαγές κέρδους είναι εξαιρετικά γρήγορες, ενώ άλλες παίρνουν δευτερόλεπτα ή και λεπτά για να σταθεροποιηθούν [48]. Συνήθως, η προσαρμογή φωτός διαρκεί περίπου 5 λεπτά (έξι φορές ταχύτερα από τη σκοτεινή προσαρμογή). Αυτό μπορεί να δείχνει την επίδραση των μεταδεκτικών περιοχών.
Το σχήμα 6 δείχνει παραδείγματα προσαρμογής φωτός [43]. Εάν χρησιμοποιήσαμε μία μόνο λειτουργία απόκρισης για να χαρτογραφήσουμε το μεγάλο εύρος εντάσεων στην έξοδο του οπτικού συστήματος, τότε θα είχαμε μόνο μια πολύ μικρή κλίμακα στη διάθεσή μας για μια δεδομένη σκηνή. Είναι σαφές ότι με μια τέτοια λειτουργία απόκρισης, η αντιληπτή αντίθεση οποιασδήποτε δεδομένης σκηνής θα ήταν περιορισμένη και η οπτική ευαισθησία στις αλλαγές θα υποβαθμιζόταν σοβαρά λόγω ζητημάτων σήματος-θορύβου. Αυτή η περίπτωση φαίνεται από τη διακεκομμένη γραμμή. Από την άλλη πλευρά, οι συμπαγείς γραμμές αντιπροσωπεύουν οικογένειες οπτικών αποκρίσεων. Αυτές οι καμπύλες καταγράφουν το εύρος χρήσιμου φωτισμού σε οποιαδήποτε δεδομένη σκηνή στο πλήρες δυναμικό εύρος της οπτικής εξόδου, με αποτέλεσμα την καλύτερη οπτική αντίληψη για κάθε κατάσταση. Η προσαρμογή φωτός μπορεί να θεωρηθεί ως η διαδικασία της ολίσθησης της καμπύλης οπτικής απόκρισης κατά μήκος του άξονα της στάθμης φωτισμού έως ότου επιτευχθεί το βέλτιστο επίπεδο για τις δεδομένες συνθήκες προβολής.
Χρωμική προσαρμογή
[επεξεργασία]Η γενική έννοια της χρωματικής προσαρμογής συνίσταται στην μεταβολή του ύψους των καμπυλών φασματικής απόκρισης των τριών κώνων. Αυτή η ρύθμιση προκύπτει επειδή η προσαρμογή φωτός συμβαίνει ανεξάρτητα σε κάθε κατηγορία κώνου. Μια συγκεκριμένη διατύπωση αυτής της υπόθεσης είναι γνωστή ως προσαρμογή von Kries. Αυτή η υπόθεση δηλώνει ότι η απόκριση προσαρμογής λαμβάνει χώρα σε κάθε έναν από τους τρεις τύπους κώνων χωριστά και ισοδυναμεί με τον πολλαπλασιασμό των σταθερών φασματικών ευαισθησιών τους με μία σταθερά κλίμακας [49]. Εάν τα βάρη κλίμακας (επίσης γνωστά ως συντελεστές von Kries) είναι αντιστρόφως ανάλογες με την απορρόφηση του φωτός από κάθε τύπο κώνου (δηλαδή χαμηλότερη απορρόφηση θα απαιτήσει μεγαλύτερο συντελεστή), τότε η κλίμακα von Kries διατηρεί μια σταθερή μέση απόκριση σε κάθε κατηγορία κώνου. Αυτό παρέχει έναν απλό αλλά ισχυρό μηχανισμό για τη διατήρηση του αντιληπτού χρώματος αντικειμένων παρά τις αλλαγές στον φωτισμό. Κάτω από μια σειρά διαφορετικών συνθηκών, η κλίμακα von Kries παρέχει μια καλή εικόνα των επιπτώσεων της προσαρμογής φωτός στην ευαισθησία και την εμφάνιση χρώματος [50], [51].
Ο ευκολότερος τρόπος για να απεικονίσετε τη χρωματική προσαρμογή είναι να εξετάσετε ένα λευκό αντικείμενο υπό διαφορετικούς τύπους φωτισμού. Για παράδειγμα, ας εξετάσουμε την εξέταση ενός τεμαχίου χαρτιού κάτω από το φως της ημέρας, φθορισμού και φωτισμού πυρακτώσεως. Το φως της ημέρας περιέχει σχετικώς πολύ περισσότερη ενέργεια μικρού μήκους κύματος από το φθορίζον φως και ο φωτισμός πυρακτώσεως περιέχει σχετικώς πολύ περισσότερη ενέργεια μακρού μήκους κύματος από το φθορίζον φως. Ωστόσο, παρά τις διαφορετικές συνθήκες φωτισμού, το χαρτί διατηρεί περίπου την λευκή του εμφάνιση κάτω από τις τρεις πηγές φωτός. Αυτό συμβαίνει επειδή το σύστημα S-cone γίνεται σχετικά λιγότερο ευαίσθητο υπό το φως της ημέρας (προκειμένου να αντισταθμιστεί η πρόσθετη ενέργεια μικρού μήκους κύματος) και το σύστημα Lκώνων καθίσταται σχετικά λιγότερο ευαίσθητο υπό τον φωτισμό πυρακτώσεως (προκειμένου να αντισταθμιστεί η πρόσθετη μακροπρόθεσμη ακτινοβολία) ενέργεια μήκους κύματος) [43].
- ↑ Conway, Bevil R (2009). "Color vision, cones, and color-coding in the cortex". The neuroscientist 15: 274-290.
- ↑ Russell, Richard and Sinha, Pawan} (2007). "Real-world face recognition: The importance of surface reflectance properties". Perception 36 (9).
- ↑ Gegenfurtner, Karl R and Rieger, Jochem (2000). "Sensory and cognitive contributions of color to the recognition of natural scenes". Current Biology 10 (13): 805-808.
- ↑ Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke (2006). "Bare skin, blood and the evolution of primate colour vision". Biology letters 2 (2): 217-221.
- ↑ Beretta, Giordano (2000). Understanding Color. Hewlett-Packard.
- ↑ 6,0 6,1 Boynton, Robert M (1988). "Color vision". Annual review of psychology 39 (1): 69-100.
- ↑ Grassmann, Hermann (1853). "Zur theorie der farbenmischung". Annalen der Physik 165 (5): 69-84.
- ↑ Konig, Arthur and Dieterici, Conrad (1886). "Die Grundempfindungen und ihre intensitats-Vertheilung im Spectrum". Koniglich Preussischen Akademie der Wissenschaften.
- ↑ Smith, Vivianne C and Pokorny, Joel (1975). "Spectral sensitivity of the foveal cone photopigments between 400 and 500 nm". Vision research 15 (2): 161-171.
- ↑ Vos, JJ and Walraven, PL (1971). "On the derivation of the foveal receptor primaries". Vision Research 11 (8): 799-818.
- ↑ 11,0 11,1 Gegenfurtner, Karl R and Kiper, Daniel C (2003). "Color vision". Neuroscience 26 (1): 181.
- ↑ Kaiser, Peter K and Boynton, Robert M (1985). "Role of the blue mechanism in wavelength discrimination". Vision research 125 (4): 523-529.
- ↑ Paulus, Walter and Kroger-Paulus, Angelika (1983). "A new concept of retinal colour coding". Vision research 23 (5): 529-540.
- ↑ Nerger, Janice L and Cicerone, Carol M (1992). "The ratio of L cones to M cones in the human parafoveal retina". Vision research 32 (5): 879-888.
- ↑ Neitz, Jay and Carroll, Joseph and Yamauchi, Yasuki and Neitz, Maureen and Williams, David R (2002). "Color perception is mediated by a plastic neural mechanism that is adjustable in adults". Neuron 35 (4): 783-792.
- ↑ Jacobs, Gerald H and Williams, Gary A and Cahill, Hugh and Nathans, Jeremy (2007). "Emergence of novel color vision in mice engineered to express a human cone photopigment". Science 315 (5819): 1723-1725.
- ↑ Osorio, D and Ruderman, DL and Cronin, TW (1998). "Estimation of errors in luminance signals encoded by primate retina resulting from sampling of natural images with red and green cones". JOSA A 15 (1): 16-22.
- ↑ Kersten, Daniel (1987). "Predictability and redundancy of natural images". JOSA A 4 (112): 2395-2400.
- ↑ Jolliffe, I. T. (2002). Principal Component Analysis. Springer.
- ↑ Buchsbaum, Gershon and Gottschalk, A (1983). "Trichromacy, opponent colours coding and optimum colour information transmission in the retina". Proceedings of the Royal society of London. Series B. Biological sciences 220 (1218): 89-113.
- ↑ Zaidi, Qasim (1997). "Decorrelation of L-and M-cone signals". JOSA A 14 (12): 3430-3431.
- ↑ Ruderman, Daniel L and Cronin, Thomas W and Chiao, Chuan-Chin (1998). "Statistics of cone responses to natural images: Implications for visual coding". JOSA A 15 (8): 2036-2045.
- ↑ Lee, BB and Martin, PR and Valberg, A (1998). "The physiological basis of heterochromatic flicker photometry demonstrated in the ganglion cells of the macaque retina". The Journal of Physiology 404 (1): 323-347.
- ↑ Derrington, Andrew M and Krauskopf, John and Lennie, Peter (1984). "Chromatic mechanisms in lateral geniculate nucleus of macaque". The Journal of Physiology 357 (1): 241-265.
- ↑ Martin, Paul R and Lee, Barry B and White, Andrew JR and Solomon, Samuel G and Ruttiger, Lukas (2001). "Chromatic sensitivity of ganglion cells in the peripheral primate retina". Nature 410 (6831): 933-936.
- ↑ Perry, VH and Oehler, R and Cowey, A (1984). "Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey". Neuroscience 12 (4): 1101--1123.